

Neutral lipids associated with haemozoin mediate efficient and rapid β-haematin formation at physiological pH, temperature and ionic composition
- PMID: 23043460
- PMCID: PMC3479076
- DOI: 10.1186/1475-2875-11-337
Neutral lipids associated with haemozoin mediate efficient and rapid β-haematin formation at physiological pH, temperature and ionic composition
Abstract
Background: The malaria parasite disposes of host-derived ferrihaem (iron(III)protoporphyrin IX, Fe(III)PPIX) by conversion to crystalline haemozoin in close association with neutral lipids. Lipids mediate synthetic haemozoin (β-haematin) formation very efficiently. However, the effect on reaction rates of concentrations of lipid, Fe(III)PPIX and physiologically relevant ions and biomolecules are unknown.
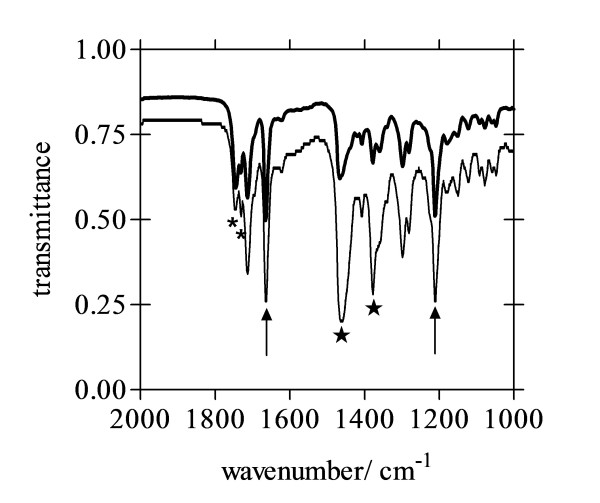
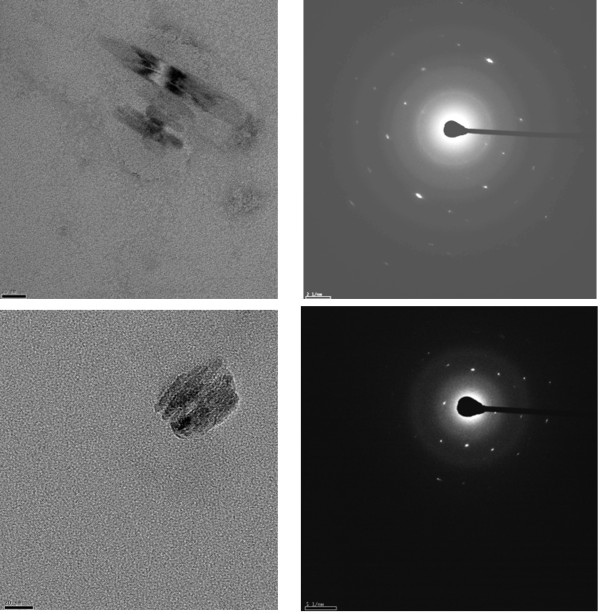
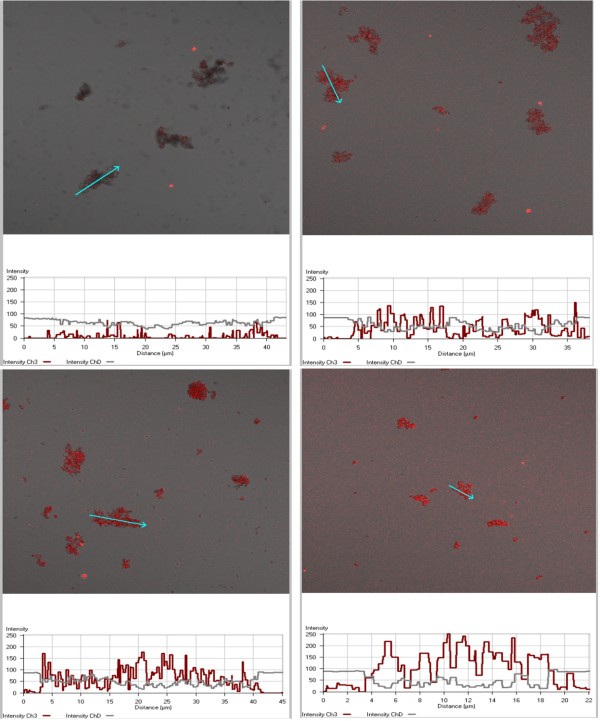
Methods: Lipid emulsions containing Fe(III)PPIX were prepared in aqueous medium (pH 4.8, 37°C) to mediate β-haematin formation. The reaction was quenched at various times and free Fe(III)PPIX measured colorimetrically as a pyridine complex and the kinetics and yields analysed. Products were also characterized by FTIR, TEM and electron diffraction. Autofluorescence was also used to monitor β-haematin formation by confocal microscopy.
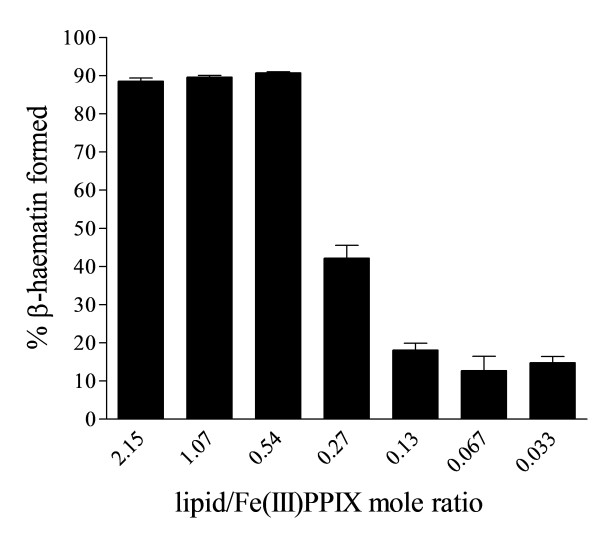
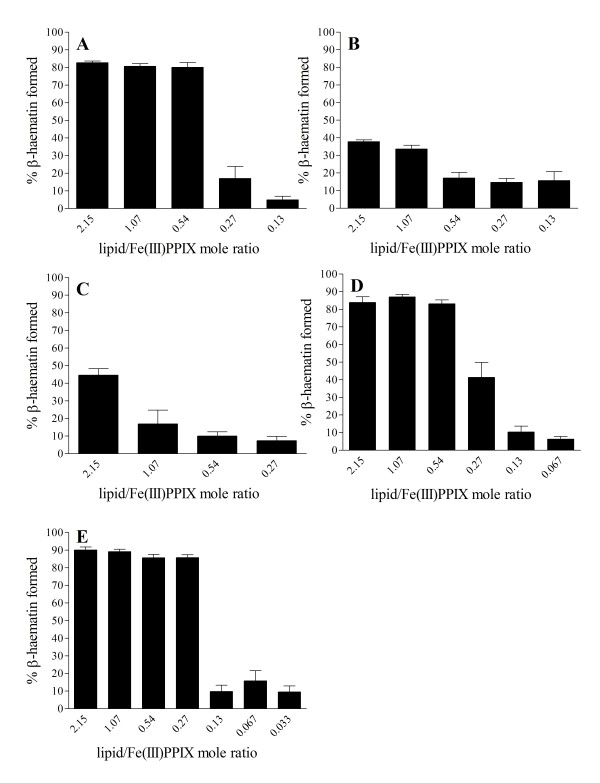
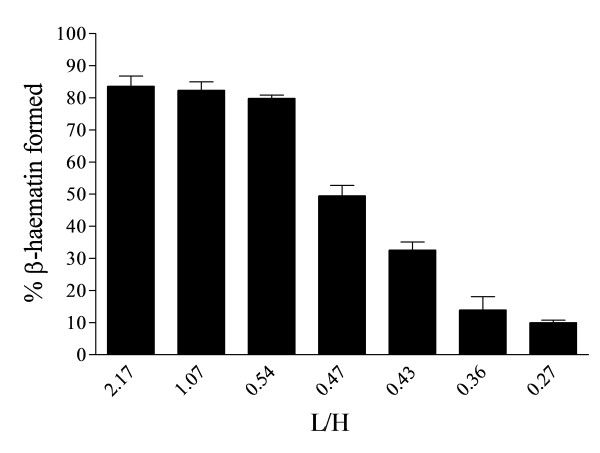
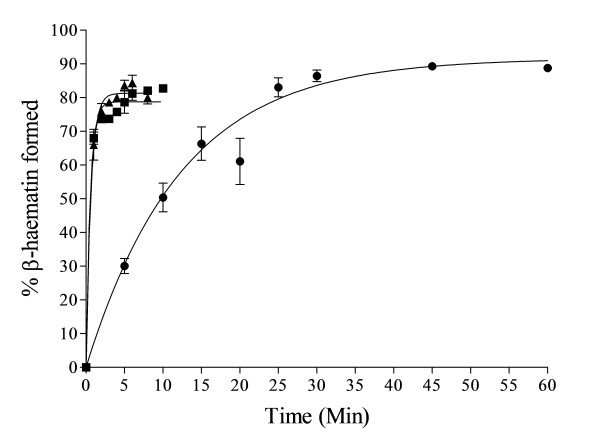
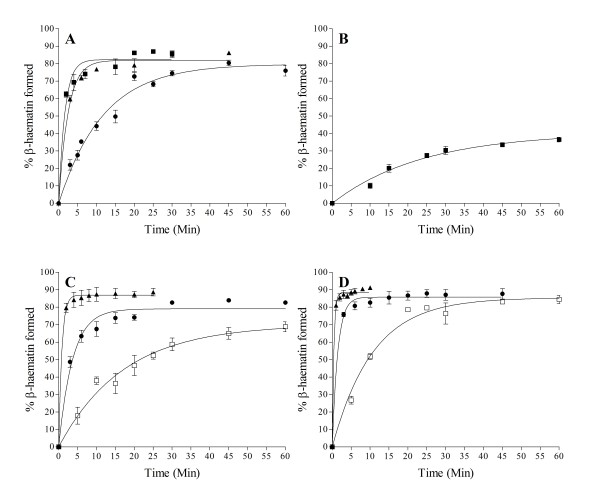
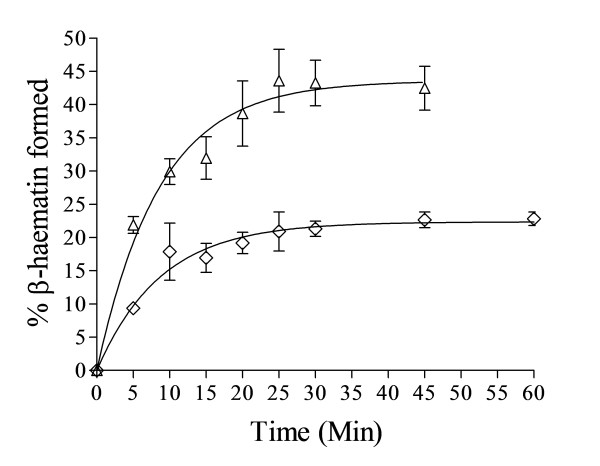
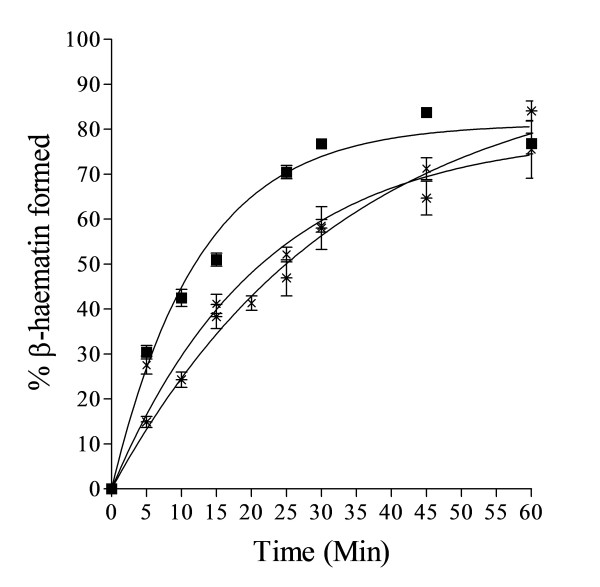
Results: At fixed Fe(III)PPIX concentration, β-haematin yields remained constant with decreasing lipid concentration until a cut-off ratio was reached whereupon efficiency decreased dramatically. For the haemozoin-associated neutral lipid blend (NLB) and monopalmitoylglycerol (MPG), this occurred below a lipid/Fe(III)PPIX (L/H) ratio of 0.54. Rate constants were found to increase with L/H ratio above the cut-off. At 16 μM MPG, Fe(III)PPIX concentration could be raised until the L/H ratio reached the same ratio before a sudden decline in yield was observed. MPG-mediated β-haematin formation was relatively insensitive to biologically relevant cations (Na(+), K(+), Mg(2+), Ca(2+)), or anions (H(2)PO(4)(-), HCO(3)(-), ATP, 2,3-diphosphoglycerate, glutathione). Confocal microscopy demonstrated β-haematin formation occurs in association with the lipid particles.
Conclusions: Kinetics of β-haematin formation have shown that haemozoin-associated neutral lipids alone are capable of mediating β-haematin formation at adequate rates under physiologically realistic conditions of ion concentrations to account for haemozoin formation.
Figures
References
-
- Tekwani BL, Walker LA. Targeting the hemozoin synthesis pathway for new antimalarial drug discovery: technologies for in vitro β-hematin formation assay. Combinatorial Chem High Througput Scr. 2005;8:61–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
