

Sirtuin-4 modulates sensitivity to induction of the mitochondrial permeability transition pore
- PMID: 23044393
- PMCID: PMC3498821
- DOI: 10.1016/j.bbabio.2012.09.016
Sirtuin-4 modulates sensitivity to induction of the mitochondrial permeability transition pore
Retraction in
-
Retraction notice to - Sirtuin-4 modulates sensitivity to induction of the mitochondrial permeability transition pore.Biochim Biophys Acta. 2016 Sep;1857(9):1641. doi: 10.1016/j.bbabio.2016.06.009. Biochim Biophys Acta. 2016. PMID: 27473497 Free PMC article. No abstract available.
Abstract
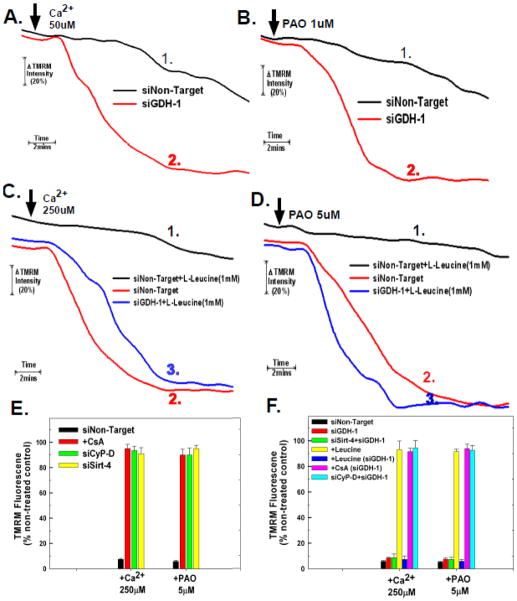
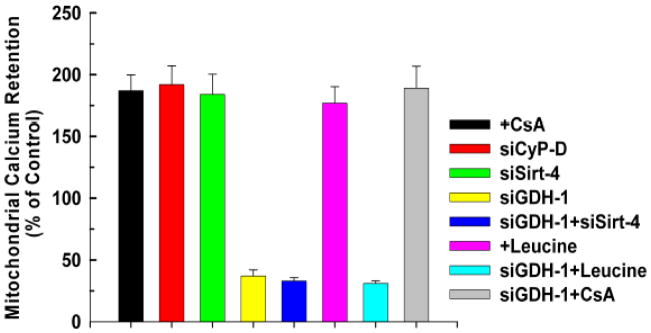
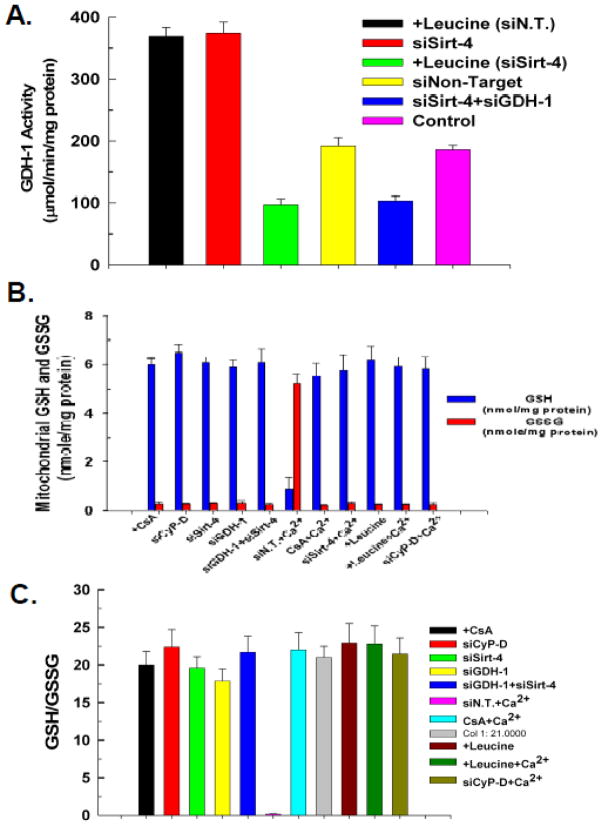
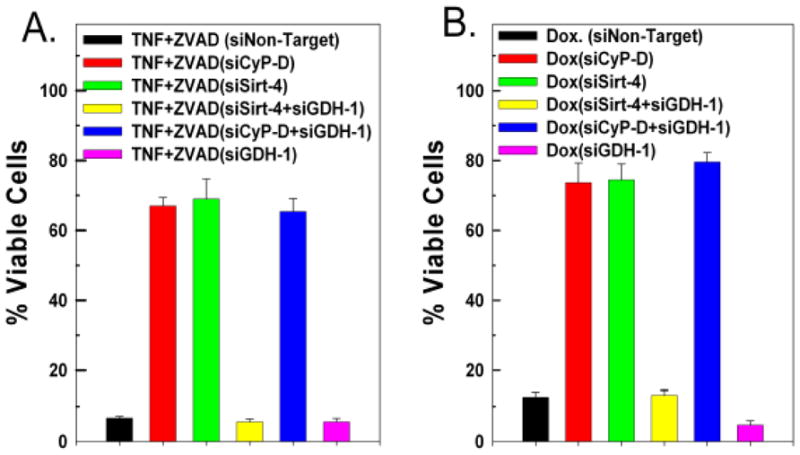
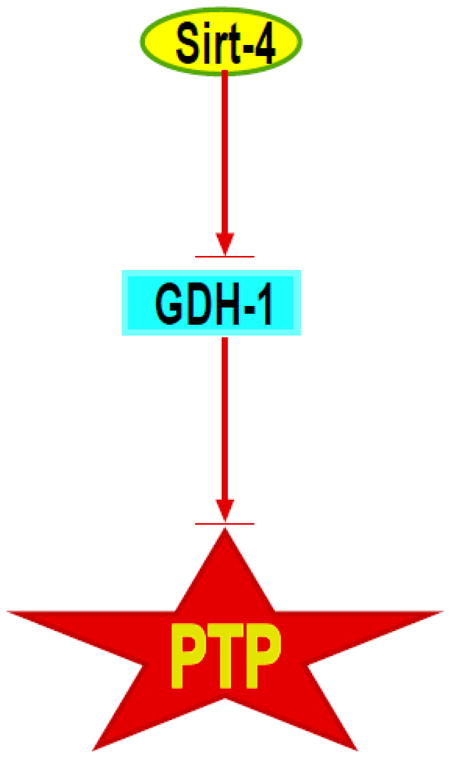
The sustained opening of the mitochondrial permeability transition pore (PTP) is a decisive event in the onset of irreversible cell injury. The PTP is modulated by numerous exogenous and endogenous effectors, including mitochondrial membrane potential, ions and metabolites. Mitochondrial sirtuins have recently emerged as pivotal mediators of mitochondrial metabolism. In the present study, we demonstrate that sirt-4 modulates sensitivity to PTP onset induced by calcium and the oxidative cross linking reagent phenylarsine oxide, and PTP dependent cytotoxicity brought about by TNF or doxorubicin. Moreover, the ability of sirt-4 to modulate onset of the PTP is dependent on the expression of glutamate dehydrogenase-1.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
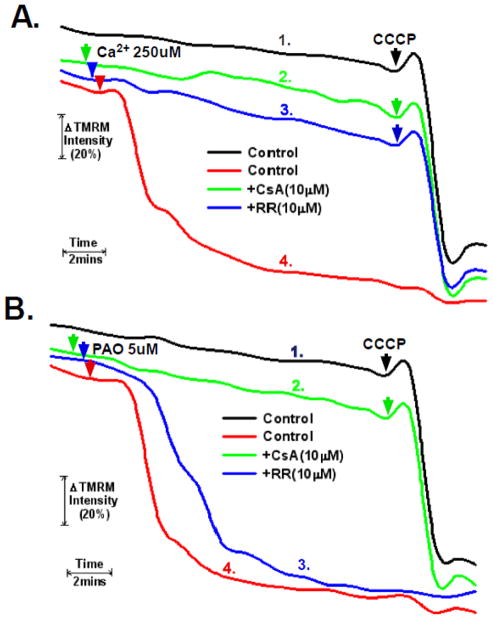
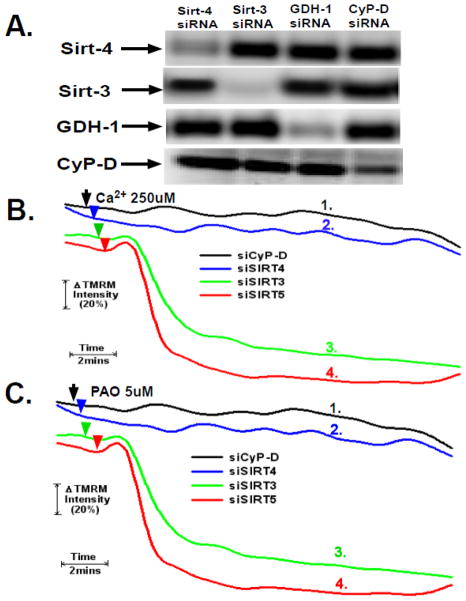
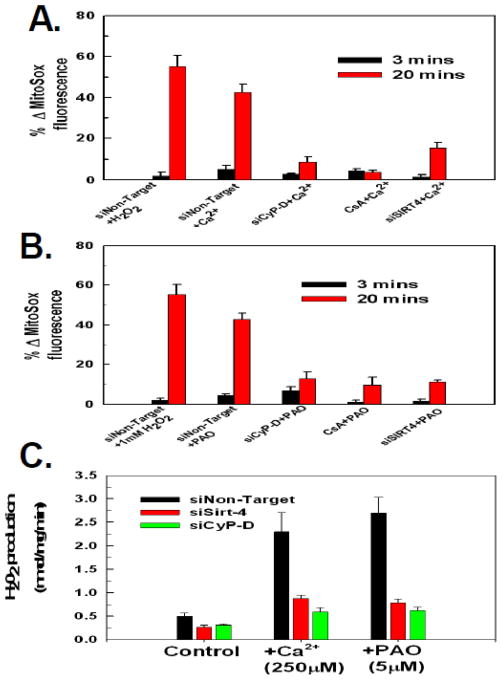
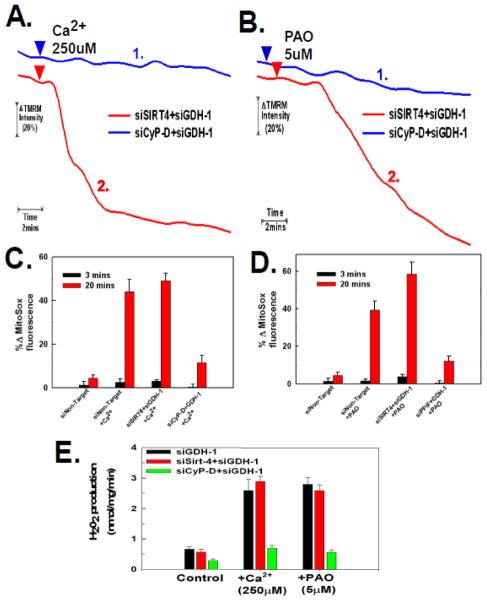
Comment in
-
Findings of research misconduct.NIH Guide Grants Contracts (Bethesda). 2016 Jun 10:NOT-OD-16-104. NIH Guide Grants Contracts (Bethesda). 2016. PMID: 27311138 Free PMC article. No abstract available.
-
Findings of Research Misconduct.Fed Regist. 2016 May 13;81(93):29871. Fed Regist. 2016. PMID: 27737277 Free PMC article. No abstract available.
References
-
- Di Lisa F, Menabo R, Canton M, Barile M, Bernardi P. Opening of the mitochondrial permeability transition pore causes depletion of mitochondrial and cytosolic NAD+ and is a causative event in the death of myocytes in postischemic reperfusion of the heart. J Biol Chem. 2001;276:2571–2575. - PubMed
-
- Halestrap AP, Connern CP, Griffiths EJ, Kerr PM. Cyclosporin A binding to mitochondrial cyclophilin inhibits the permeability transition pore and protects hearts from ischaemia/reperfusion injury. Molecular & Cellular Biochemistry. 1997;174:167–172. - PubMed
-
- Baines CP, Kaiser RA, Purcell NH, Blair NS, Osinska H, Hambleton MA, Brunskill EW, Sayen MR, Gottlieb RA, Dorn GW, Robbins J, Molkentin JD. Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature. 2005;434:658–662. - PubMed
-
- Nakagawa T, Shimizu S, Watanabe T, Yamaguchi O, Otsu K, Yamagata H, Inohara H, Kubo T, Tsujimoto Y. Cyclophilin D-dependent mitochondrial permeability transition regulates some necrotic but not apoptotic cell death. Nature. 2005;434:652–658. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
