

Anti-tumorigenic effects of Type 1 interferon are subdued by integrated stress responses
- PMID: 23045272
- PMCID: PMC3766494
- DOI: 10.1038/onc.2012.439
Anti-tumorigenic effects of Type 1 interferon are subdued by integrated stress responses
Abstract
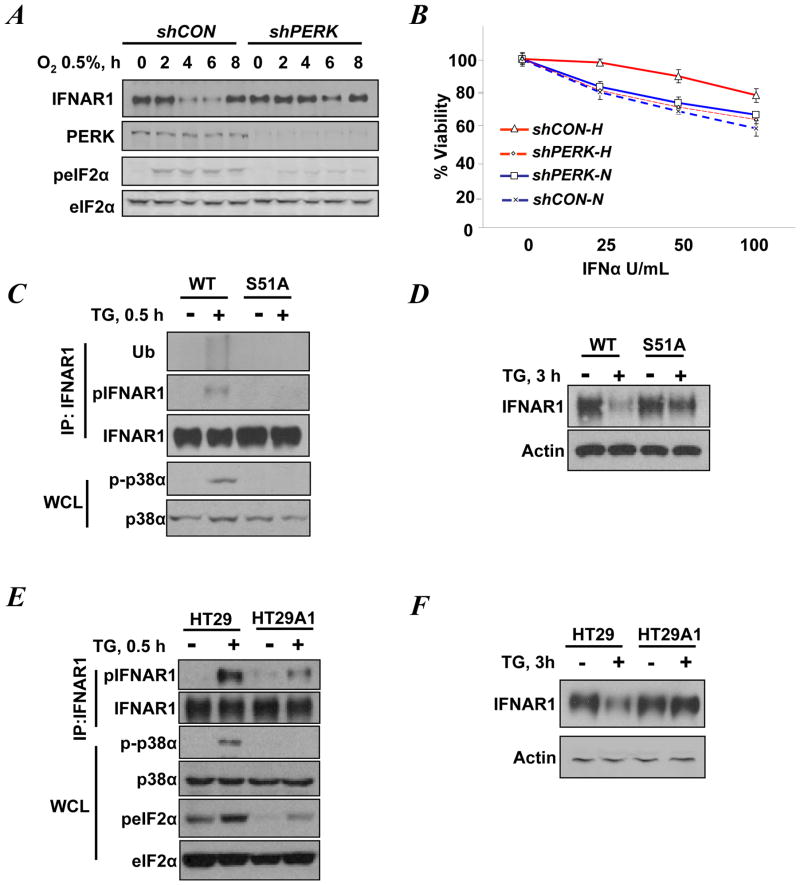
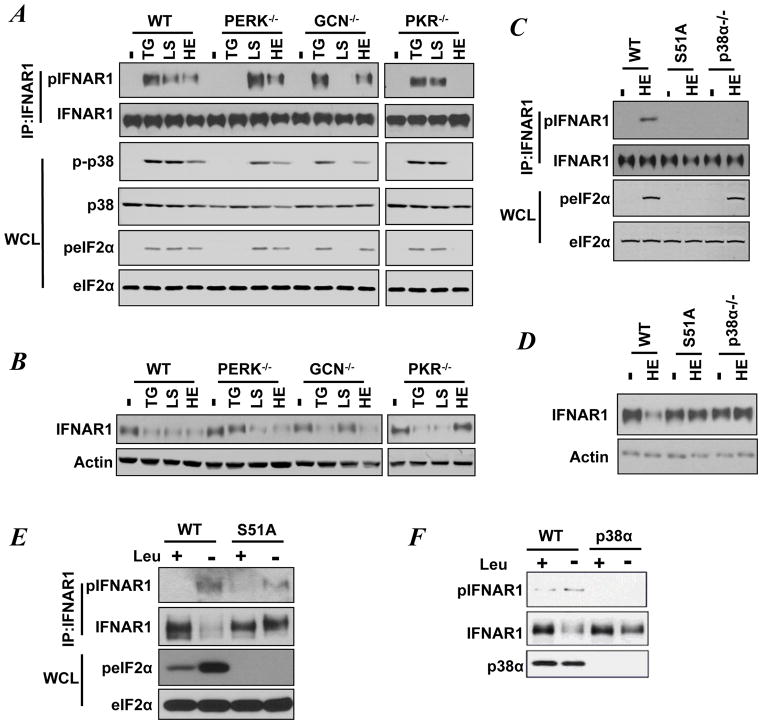
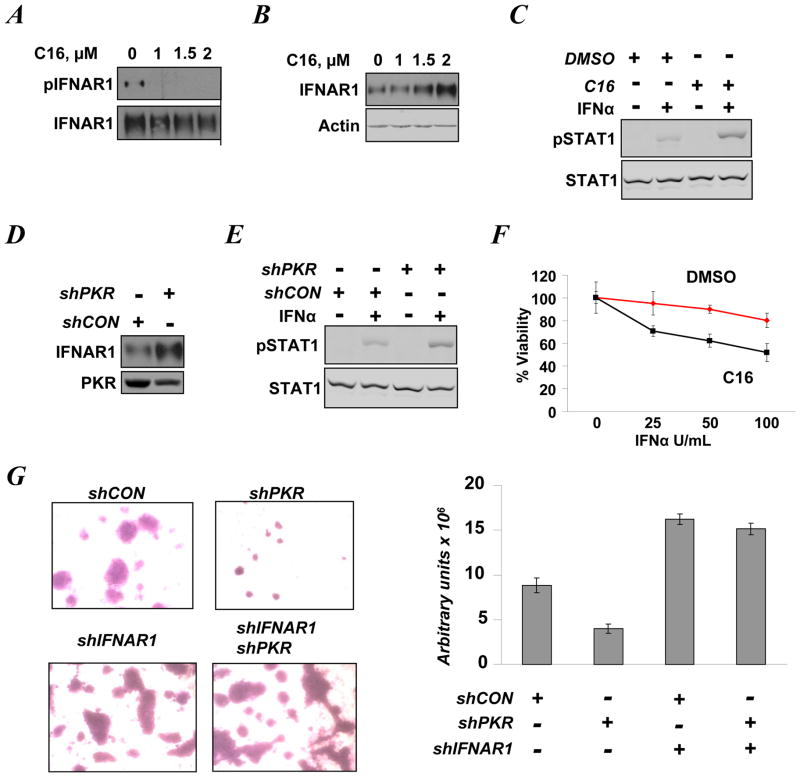
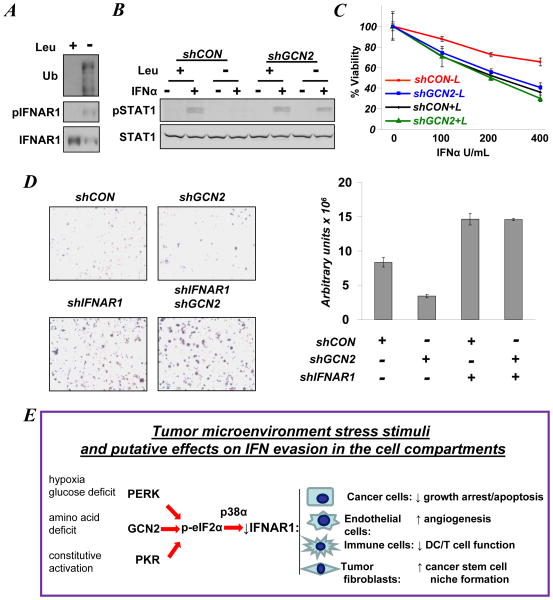
Viral and pharmacological inducers of protein kinase RNA-activated (PKR)-like ER kinase (PERK) were shown to accelerate the phosphorylation-dependent degradation of the IFNAR1 chain of the Type 1 interferon (IFN) receptor and to limit cell sensitivity to IFN. Here we report that hypoxia can elicit these effects in a PERK-dependent manner. The altered fate of IFNAR1 affected by signaling downstream of PERK depends on phosphorylation of eIF2α (eukaryotic translational initiation factor 2-α) and ensuing activation of p38α kinase. Activators of other eIF2α kinases such as PKR or GCN2 (general control nonrepressed-2) are also capable of eliminating IFNAR1 and blunting IFN responses. Modulation of constitutive PKR activity in human breast cancer cells stabilizes IFNAR1 and sensitizes these cells to IFNAR1-dependent anti-tumorigenic effects. Although downregulation of IFNAR1 and impaired IFNAR1 signaling can be elicited in response to amino-acid deficit, the knockdown of GCN2 in melanoma cells reverses these phenotypes. We propose that, in cancer cells and the tumor microenvironment, activation of diverse eIF2α kinases followed by IFNAR1 downregulation enables multiple cellular components of tumor tissue to evade the direct and indirect anti-tumorigenic effects of Type 1 IFN.
Conflict of interest statement
The authors declare no conflict of interest except for Dr. D.P. Baker who is an employee of BiogenIDEC, Inc and owns stock of this company.
Figures
References
-
- Ye J, Koumenis C. ATF4, an ER stress and hypoxia-inducible transcription factor and its potential role in hypoxia tolerance and tumorigenesis. Curr Mol Med. 2009;9(4):411–6. - PubMed
-
- Schreiber RD, Old LJ, Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011;331(6024):1565–70. - PubMed
-
- Moenner M, Pluquet O, Bouchecareilh M, Chevet E. Integrated endoplasmic reticulum stress responses in cancer. Cancer Res. 2007;67(22):10631–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
