

Distant activation of transcription: mechanisms of enhancer action
- PMID: 23045397
- PMCID: PMC3510544
- DOI: 10.1128/MCB.01127-12
Distant activation of transcription: mechanisms of enhancer action
Abstract
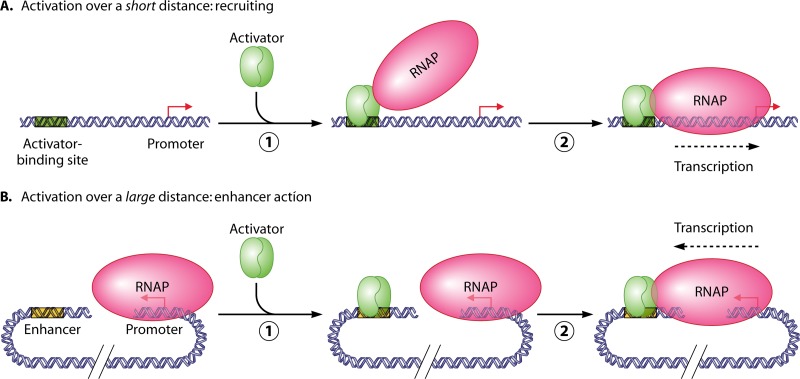
Enhancers are regulatory DNA sequences that activate transcription over long distances. Recent studies revealed a widespread role of distant activation in eukaryotic gene regulation and in development of various human diseases, including cancer. Genomic and gene-targeted studies of enhancer action revealed novel mechanisms of transcriptional activation over a distance. They include formation of stable, inactive DNA-protein complexes at the enhancer and target promoter before activation, facilitated distant communication by looping of the spacer chromatin-covered DNA, and promoter activation by mechanisms that are different from classic recruiting. These studies suggest the similarity between the looping mechanisms involved in enhancer action on DNA in bacteria and in chromatin of higher organisms.
Figures


References
-
- Bellomy GR, Record MT., Jr 1990. Stable DNA loops in vivo and in vitro: roles in gene regulation at a distance and in biophysical characterization of DNA. Prog. Nucleic Acid Res. Mol. Biol. 39:81–128 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources