

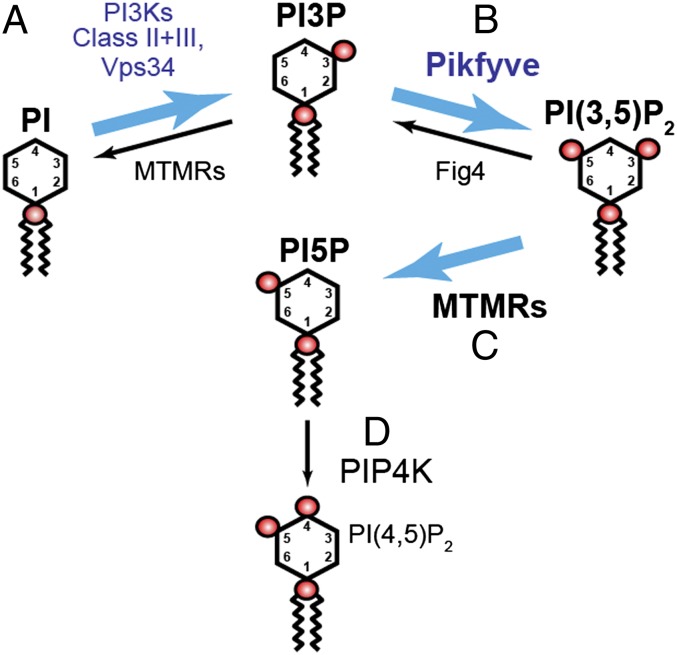
In vivo, Pikfyve generates PI(3,5)P2, which serves as both a signaling lipid and the major precursor for PI5P
- PMID: 23047693
- PMCID: PMC3491506
- DOI: 10.1073/pnas.1203106109
In vivo, Pikfyve generates PI(3,5)P2, which serves as both a signaling lipid and the major precursor for PI5P
Abstract
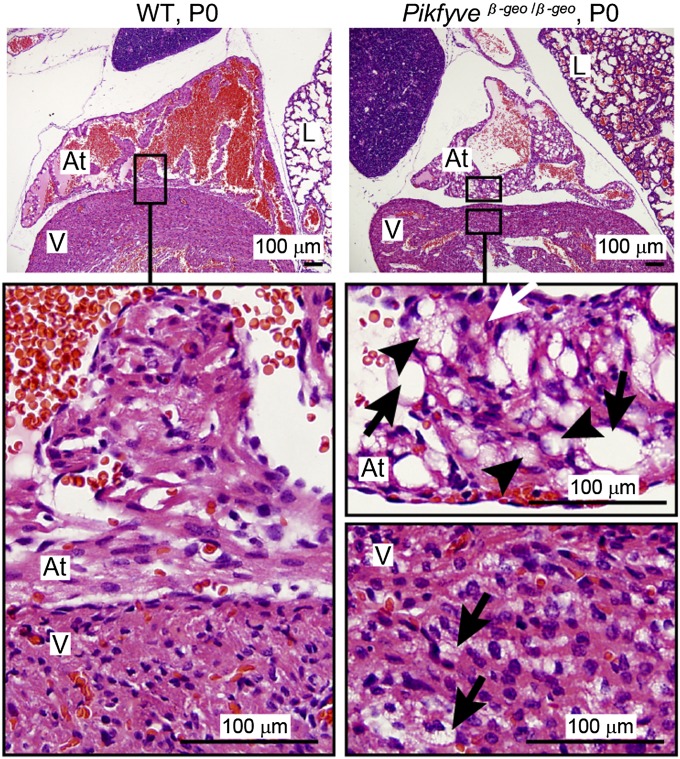
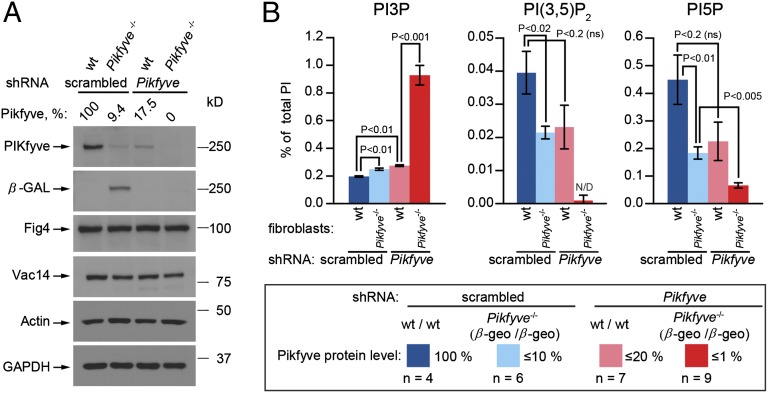
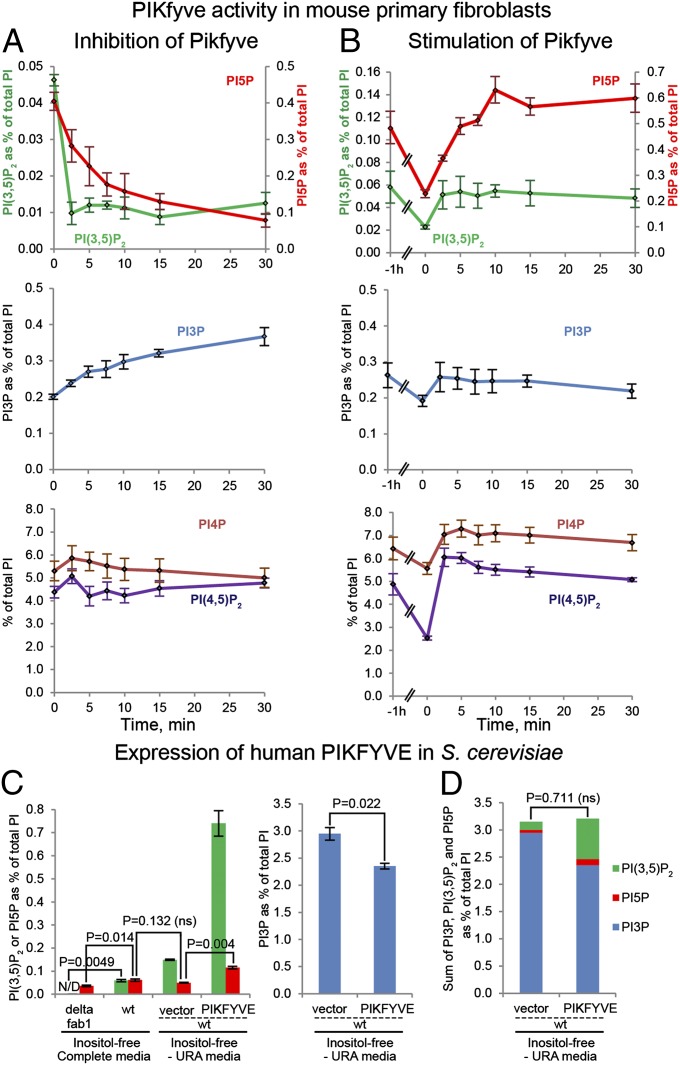
Mutations that cause defects in levels of the signaling lipid phosphatidylinositol 3,5-bisphosphate [PI(3,5)P(2)] lead to profound neurodegeneration in mice. Moreover, mutations in human FIG4 predicted to lower PI(3,5)P(2) levels underlie Charcot-Marie-Tooth type 4J neuropathy and are present in selected cases of amyotrophic lateral sclerosis. In yeast and mammals, PI(3,5)P(2) is generated by a protein complex that includes the lipid kinase Fab1/Pikfyve, the scaffolding protein Vac14, and the lipid phosphatase Fig4. Fibroblasts cultured from Vac14(-/-) and Fig4(-/-) mouse mutants have a 50% reduction in the levels of PI(3,5)P(2), suggesting that there may be PIKfyve-independent pathways that generate this lipid. Here, we characterize a Pikfyve gene-trap mouse (Pikfyve(β-geo/β-geo)), a hypomorph with ~10% of the normal level of Pikfyve protein. shRNA silencing of the residual Pikfyve transcript in fibroblasts demonstrated that Pikfyve is required to generate all of the PI(3,5)P(2) pool. Surprisingly, Pikfyve also is responsible for nearly all of the phosphatidylinositol-5-phosphate (PI5P) pool. We show that PI5P is generated directly from PI(3,5)P(2), likely via 3'-phosphatase activity. Analysis of tissues from the Pikfyve(β-geo/β-geo) mouse mutants reveals that Pikfyve is critical in neural tissues, heart, lung, kidney, thymus, and spleen. Thus, PI(3,5)P(2) and PI5P have major roles in multiple organs. Understanding the regulation of these lipids may provide insights into therapies for multiple diseases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Cooke FT, et al. The stress-activated phosphatidylinositol 3-phosphate 5-kinase Fab1p is essential for vacuole function in S. cerevisiae. Curr Biol. 1998;8(22):1219–1222. - PubMed
-
- Dove SK, et al. Vac14 controls PtdIns(3,5)P(2) synthesis and Fab1-dependent protein trafficking to the multivesicular body. Curr Biol. 2002;12(11):885–893. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
