

Functional characterization of pulmonary neuroendocrine cells in lung development, injury, and tumorigenesis
- PMID: 23047698
- PMCID: PMC3491514
- DOI: 10.1073/pnas.1207238109
Functional characterization of pulmonary neuroendocrine cells in lung development, injury, and tumorigenesis
Abstract
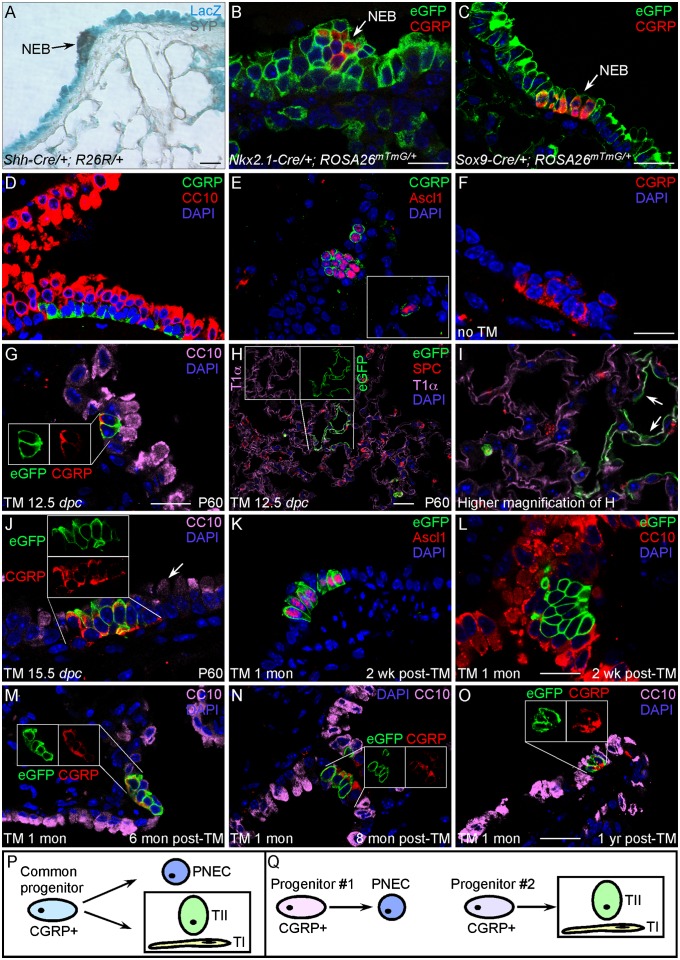
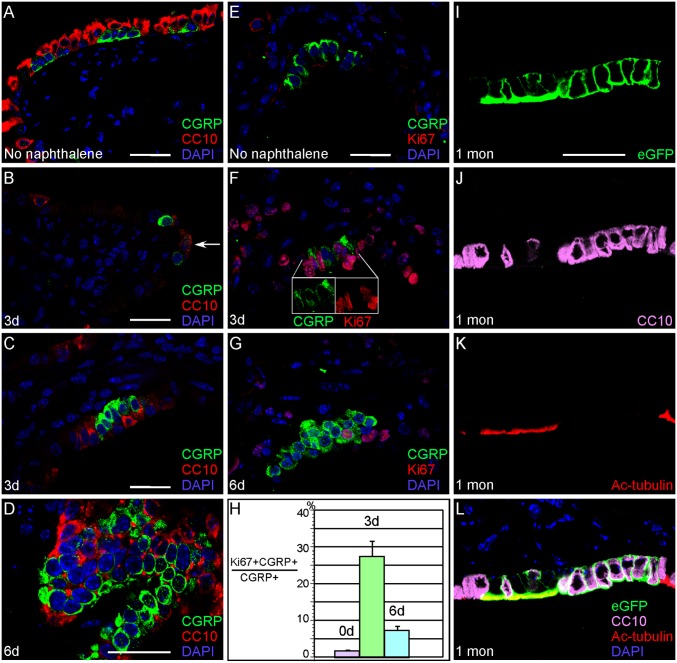
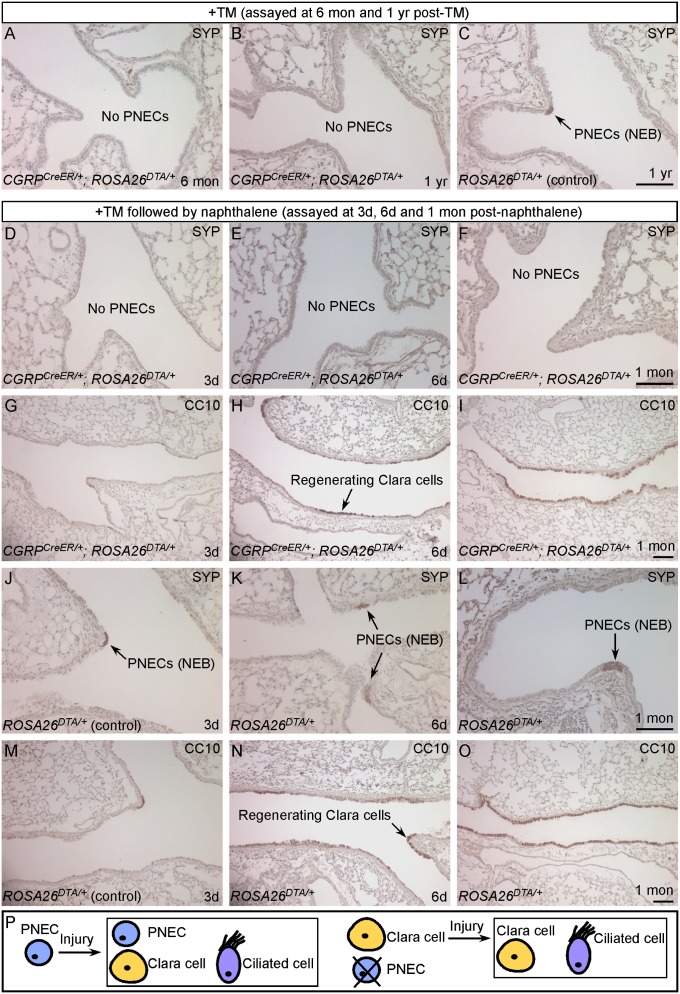
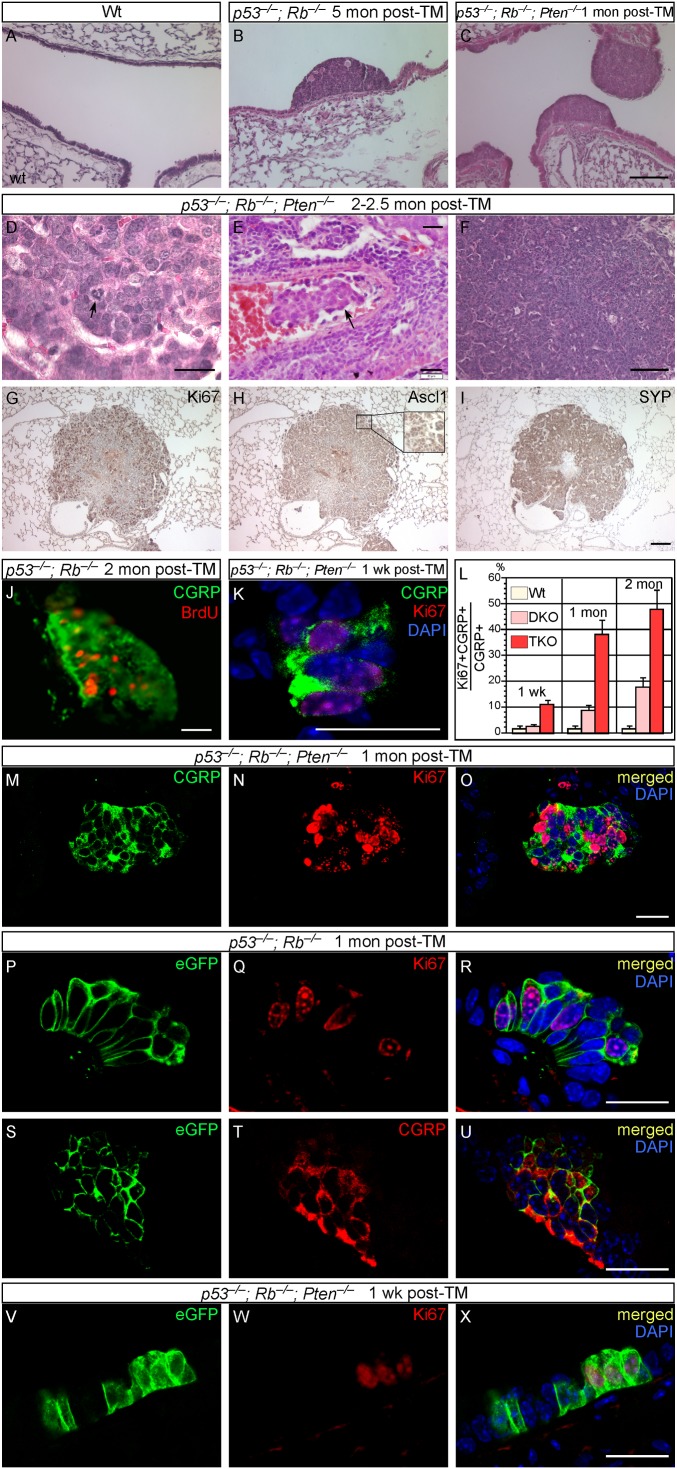
Pulmonary neuroendocrine cells (PNECs) are proposed to be the first specialized cell type to appear in the lung, but their ontogeny remains obscure. Although studies of PNECs have suggested their involvement in a number of lung functions, neither their in vivo significance nor the molecular mechanisms underlying them have been elucidated. Importantly, PNECs have long been speculated to constitute the cells of origin of human small-cell lung cancer (SCLC) and recent mouse models support this hypothesis. However, a genetic system that permits tracing the early events of PNEC transformation has not been available. To address these key issues, we developed a genetic tool in mice by introducing a fusion protein of Cre recombinase and estrogen receptor (CreER) into the calcitonin gene-related peptide (CGRP) locus that encodes a major peptide in PNECs. The CGRP(CreER) mouse line has enabled us to manipulate gene activity in PNECs. Lineage tracing using this tool revealed the plasticity of PNECs. PNECs can be colabeled with alveolar cells during lung development, and following lung injury, PNECs can contribute to Clara cells and ciliated cells. Contrary to the current model, we observed that elimination of PNECs has no apparent consequence on Clara cell recovery. We also created mouse models of SCLC in which CGRP(CreER) was used to ablate multiple tumor suppressors in PNECs that were simultaneously labeled for following their fate. Our findings suggest that SCLC can originate from differentiated PNECs. Together, these studies provide unique insight into PNEC lineage and function and establish the foundation of investigating how PNECs contribute to lung homeostasis, injury/repair, and tumorigenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Linnoila RI. Functional facets of the pulmonary neuroendocrine system. Lab Invest. 2006;86(5):425–444. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
