

The VP3 factor from viruses of Birnaviridae family suppresses RNA silencing by binding both long and small RNA duplexes
- PMID: 23049903
- PMCID: PMC3458112
- DOI: 10.1371/journal.pone.0045957
The VP3 factor from viruses of Birnaviridae family suppresses RNA silencing by binding both long and small RNA duplexes
Abstract
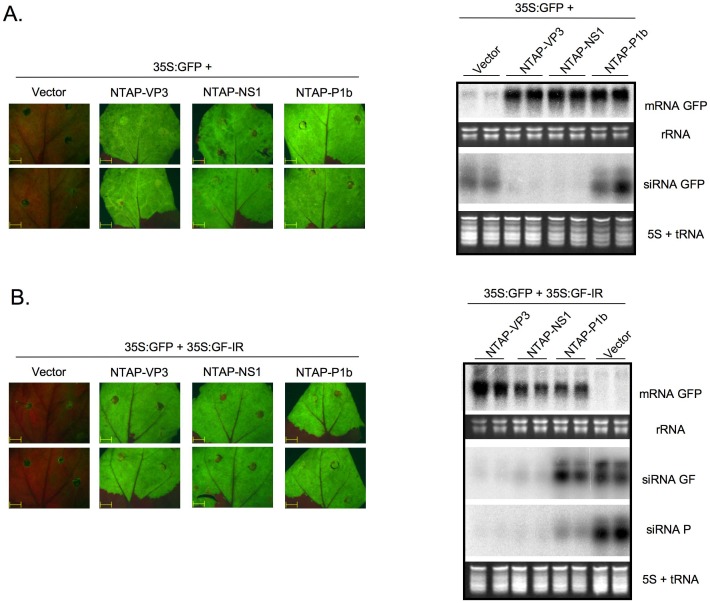
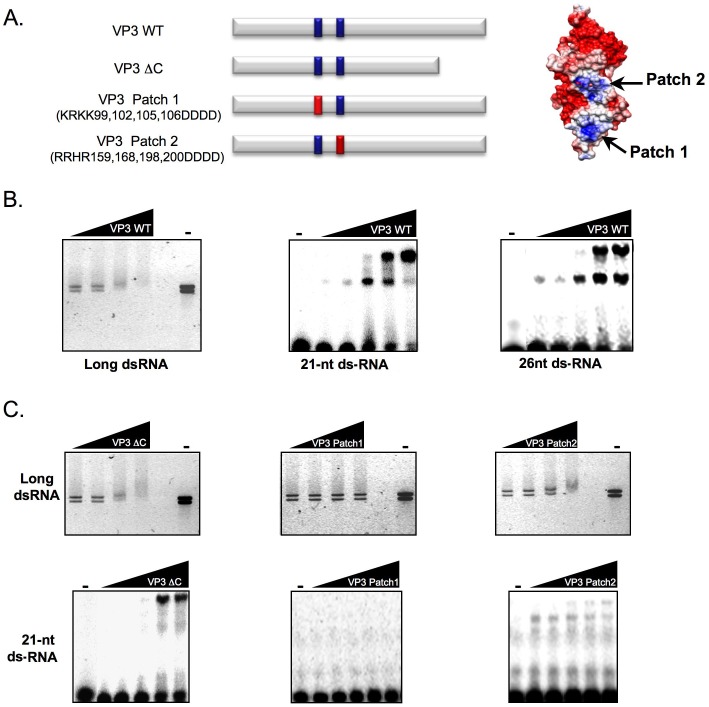
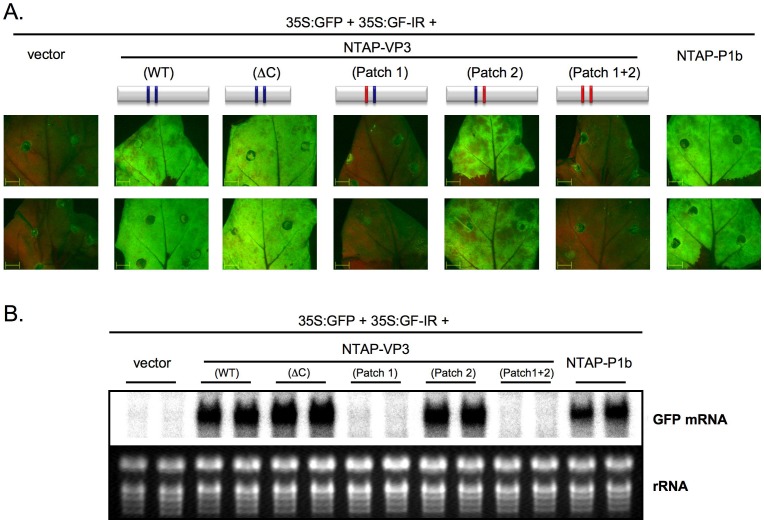
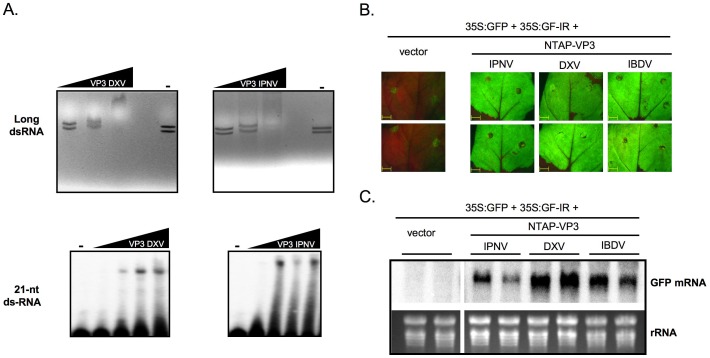
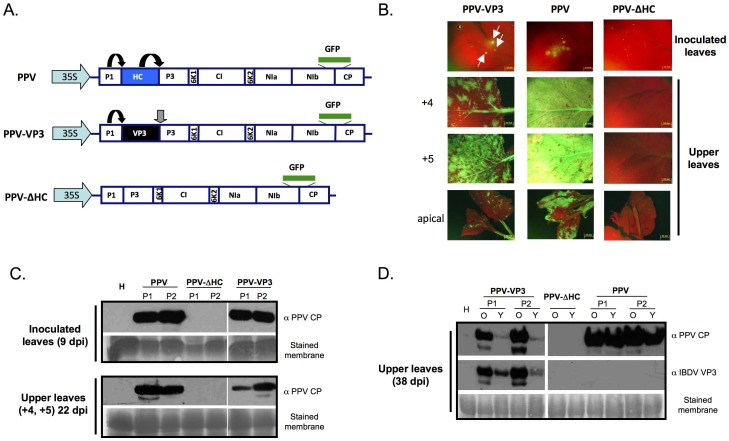
RNA silencing is directly involved in antiviral defense in a wide variety of eukaryotic organisms, including plants, fungi, invertebrates, and presumably vertebrate animals. The study of RNA silencing-mediated antiviral defences in vertebrates is hampered by the overlap with other antiviral mechanisms; thus, heterologous systems are often used to study the interplay between RNA silencing and vertebrate-infecting viruses. In this report we show that the VP3 protein of the avian birnavirus Infectious bursal disease virus (IBDV) displays, in addition to its capacity to bind long double-stranded RNA, the ability to interact with double-stranded small RNA molecules. We also demonstrate that IBDV VP3 prevents the silencing mediated degradation of a reporter mRNA, and that this silencing suppression activity depends on its RNA binding ability. Furthermore, we find that the anti-silencing activity of IBDV VP3 is shared with the homologous proteins expressed by both insect- and fish-infecting birnaviruses. Finally, we show that IBDV VP3 can functionally replace the well-characterized HCPro silencing suppressor of Plum pox virus, a potyvirus that is unable to infect plants in the absence of an active silencing suppressor. Altogether, our results support the idea that VP3 protects the viral genome from host sentinels, including those of the RNA silencing machinery.
Conflict of interest statement
Figures
Similar articles
-
Infectious Bursal Disease Virus Hijacks Endosomal Membranes as the Scaffolding Structure for Viral Replication.J Virol. 2018 May 14;92(11):e01964-17. doi: 10.1128/JVI.01964-17. Print 2018 Jun 1. J Virol. 2018. PMID: 29540593 Free PMC article.
-
VP1 and VP3 Are Required and Sufficient for Translation Initiation of Uncapped Infectious Bursal Disease Virus Genomic Double-Stranded RNA.J Virol. 2018 Jan 2;92(2):e01345-17. doi: 10.1128/JVI.01345-17. Print 2018 Jan 15. J Virol. 2018. PMID: 29093101 Free PMC article.
-
Phosphatidylinositol 3-Phosphate Mediates the Establishment of Infectious Bursal Disease Virus Replication Complexes in Association with Early Endosomes.J Virol. 2021 Feb 24;95(6):e02313-20. doi: 10.1128/JVI.02313-20. Print 2021 Feb 24. J Virol. 2021. PMID: 33361427 Free PMC article.
-
[Interaction mechanism of marine birnavirus (MABV) in fish cell lines].Uirusu. 2005 Jun;55(1):133-44. doi: 10.2222/jsv.55.133. Uirusu. 2005. PMID: 16308540 Review. Japanese.
-
Dicing and slicing: the core machinery of the RNA interference pathway.FEBS Lett. 2005 Oct 31;579(26):5822-9. doi: 10.1016/j.febslet.2005.08.079. Epub 2005 Sep 27. FEBS Lett. 2005. PMID: 16214139 Review.
Cited by
-
Mechanisms of Cell Entry by dsRNA Viruses: Insights for Efficient Delivery of dsRNA and Tools for Improved RNAi-Based Pest Control.Front Physiol. 2021 Nov 11;12:749387. doi: 10.3389/fphys.2021.749387. eCollection 2021. Front Physiol. 2021. PMID: 34858204 Free PMC article. Review.
-
Infectious Bursal Disease Virus Hijacks Endosomal Membranes as the Scaffolding Structure for Viral Replication.J Virol. 2018 May 14;92(11):e01964-17. doi: 10.1128/JVI.01964-17. Print 2018 Jun 1. J Virol. 2018. PMID: 29540593 Free PMC article.
-
Generation of recombinant VP3 protein of infectious bursal disease virus in three different expression systems, antigenic analysis of the obtained polypeptides and development of an ELISA test.Arch Virol. 2020 Jul;165(7):1611-1620. doi: 10.1007/s00705-020-04650-2. Epub 2020 May 13. Arch Virol. 2020. PMID: 32405826
-
Infectious Bursal Disease Virus VP3 Upregulates VP1-Mediated RNA-Dependent RNA Replication.J Virol. 2015 Nov;89(21):11165-8. doi: 10.1128/JVI.00218-15. Epub 2015 Aug 26. J Virol. 2015. PMID: 26311889 Free PMC article.
-
Structure and dsRNA-binding activity of the Birnavirus Drosophila X Virus VP3 protein.J Virol. 2021 Feb 15;95(4):e02166-20. doi: 10.1128/JVI.02166-20. Epub 2020 Nov 25. J Virol. 2021. PMID: 33239452 Free PMC article.
References
-
- Novina CD, Sharp PA (2004) The RNAi revolution. Nature 430: 161–164. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
