

Transient B12-dependent methyltransferase complexes revealed by small-angle X-ray scattering
- PMID: 23051056
- PMCID: PMC3484714
- DOI: 10.1021/ja3055782
Transient B12-dependent methyltransferase complexes revealed by small-angle X-ray scattering
Abstract
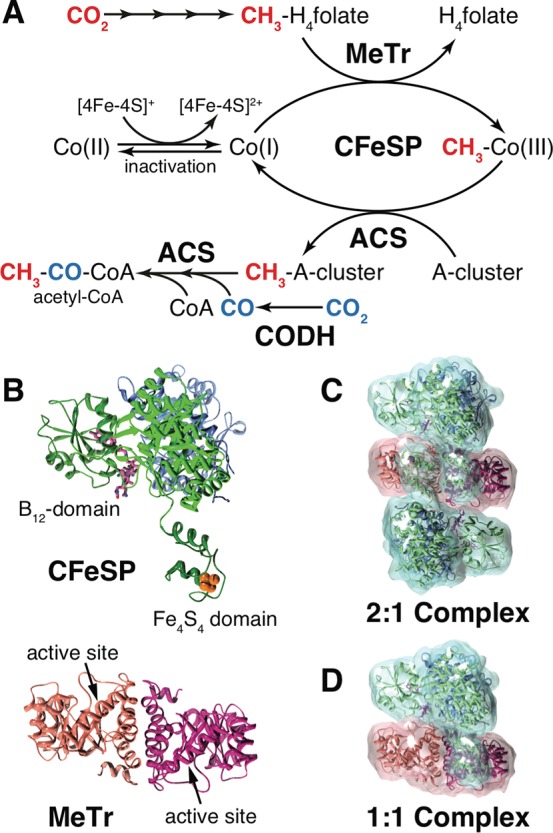
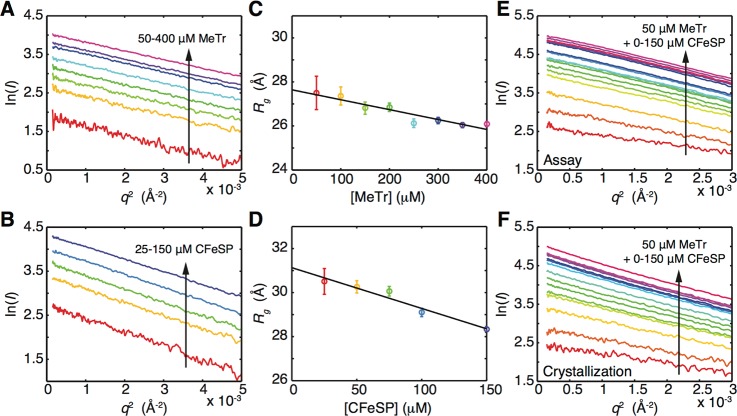
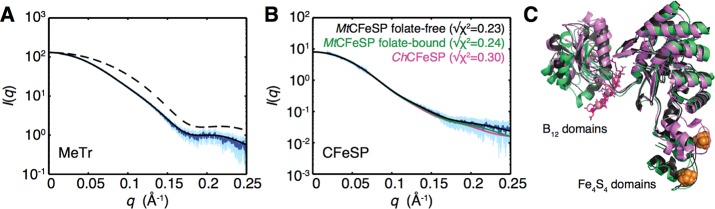
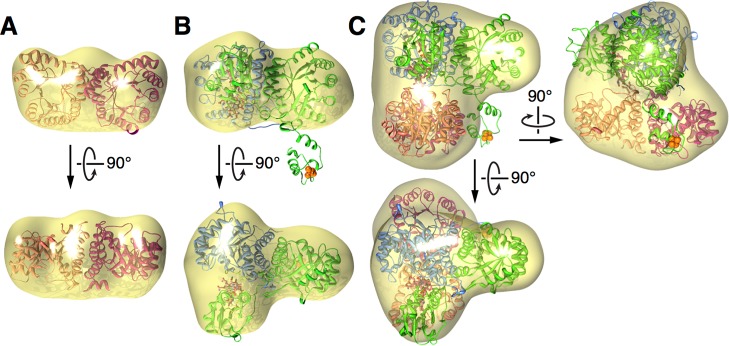
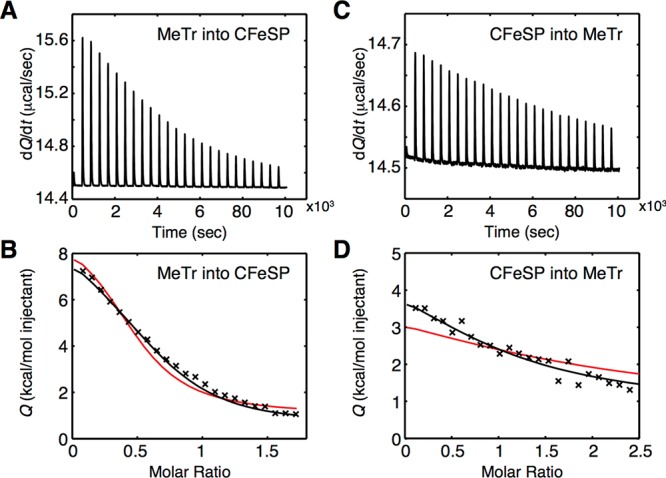
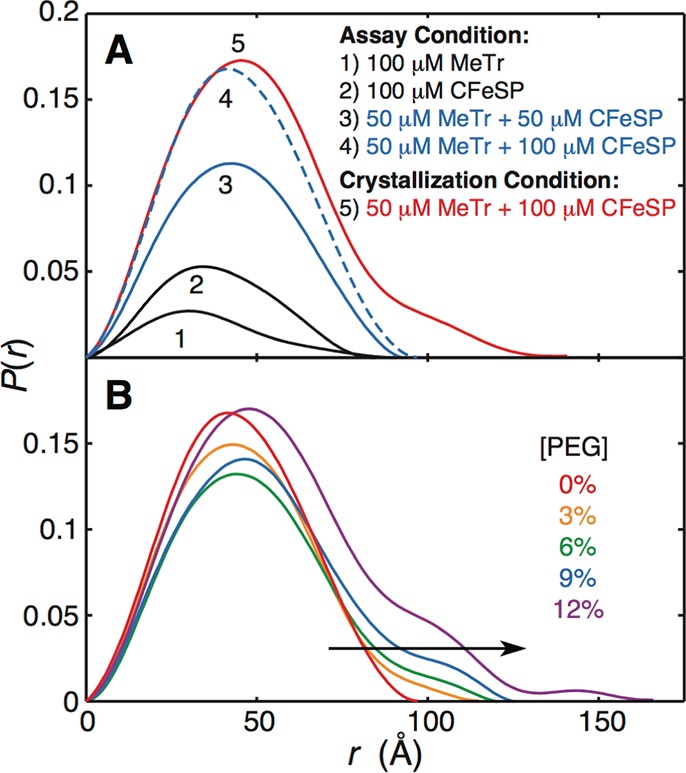
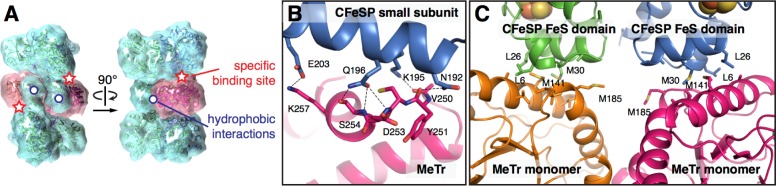
In the Wood-Ljungdahl carbon fixation pathway, protein-protein interactions between methyltransferase (MeTr) and corrinoid iron-sulfur protein (CFeSP) are required for the transfer of a methyl group. While crystal structures have been determined for MeTr and CFeSP both free and in complex, solution structures have not been established. Here, we examine the transient interactions between MeTr and CFeSP in solution using anaerobic small-angle X-ray scattering (SAXS) and present a global analysis approach for the deconvolution of heterogeneous mixtures formed by weakly interacting proteins. We further support this SAXS analysis with complementary results obtained by anaerobic isothermal titration calorimetry. Our results indicate that solution conditions affect the cooperativity with which CFeSP binds to MeTr, resulting in two distinct CFeSP/MeTr complexes with differing oligomeric compositions, both of which are active. One assembly resembles the CFeSP/MeTr complex observed crystallographically with 2:1 protein stoichiometry, while the other best fits a 1:1 CFeSP/MeTr arrangement. These results demonstrate the value of SAXS in uncovering the rich solution behavior of transient protein interactions visualized by crystallography.
Figures
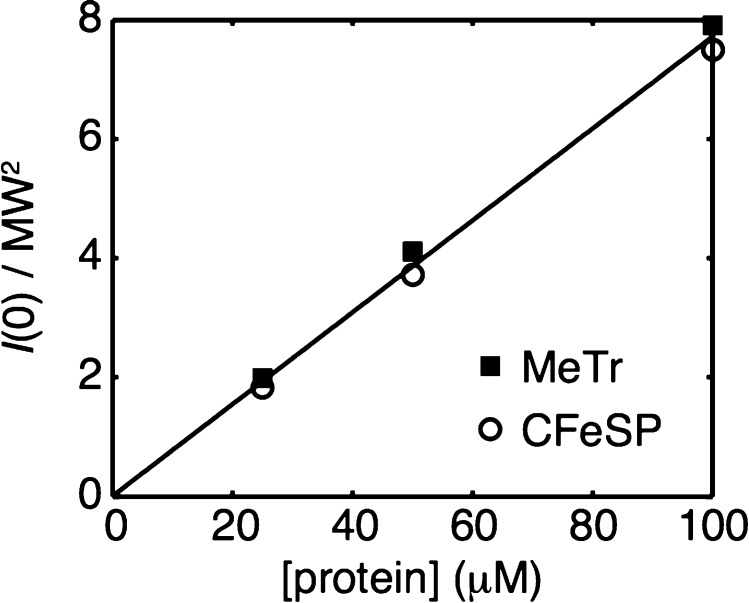
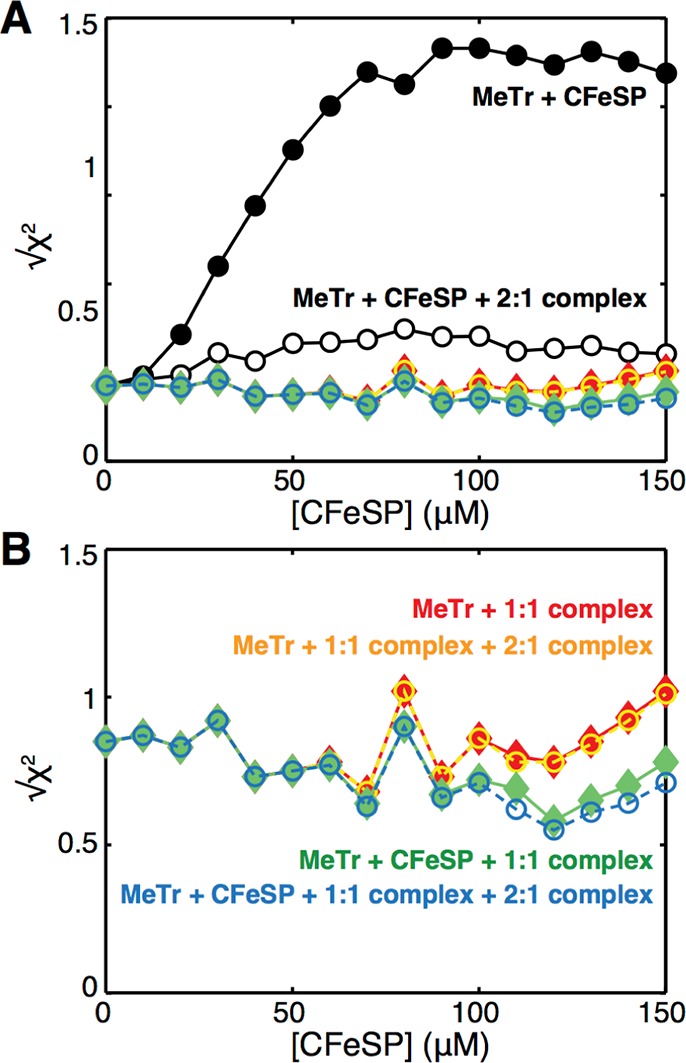
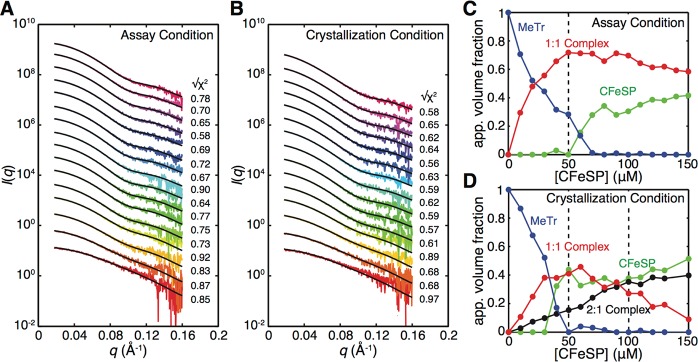
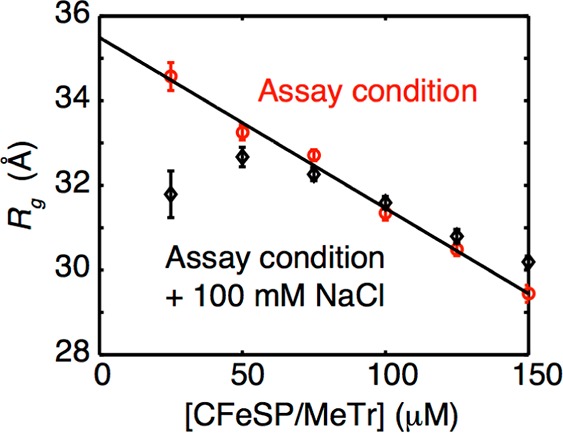
References
-
- Drake H. L.; Gössner A. S.; Daniel S. L. Ann. N.Y. Acad. Sci. 2008, 1125, 100. - PubMed
-
- Phillips J. R.; Clausen E. C.; Gaddy J. L. Appl. Biochem. Biotechnol. 1994, 45, 145.
-
- Slivka R. M.; Chinn M. S.; Grunden A. M. Biofuels 2011, 2, 405.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
