

Flavoenzymes: versatile catalysts in biosynthetic pathways
- PMID: 23051833
- PMCID: PMC3518583
- DOI: 10.1039/c2np20069d
Flavoenzymes: versatile catalysts in biosynthetic pathways
Abstract
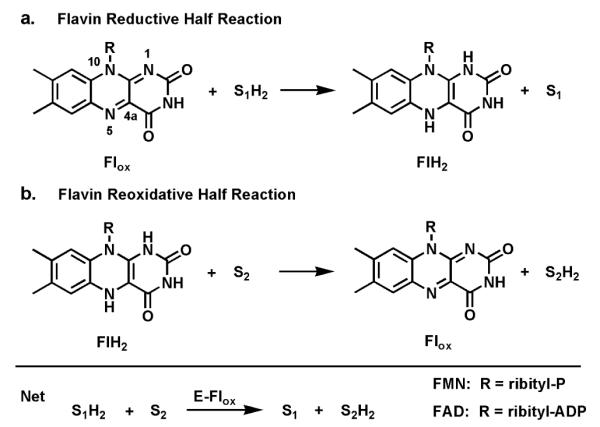
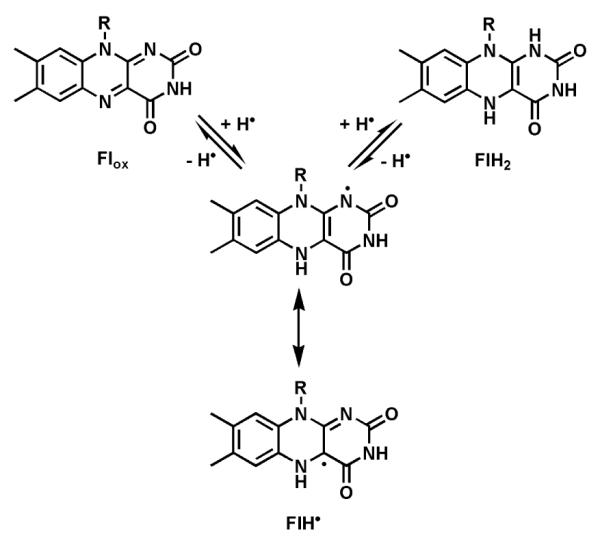
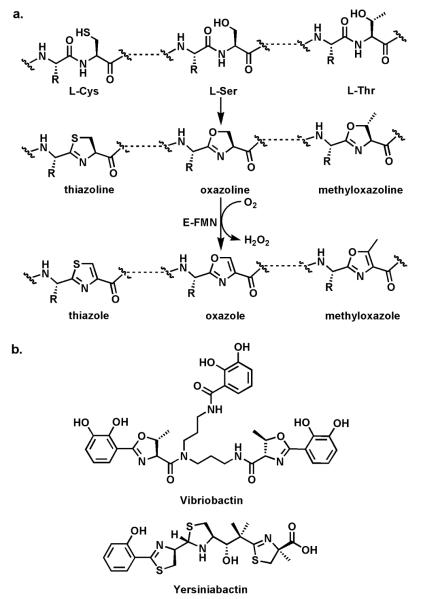
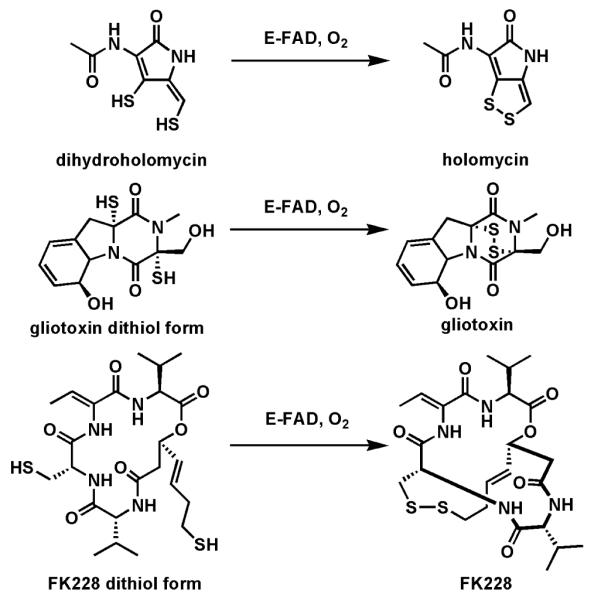
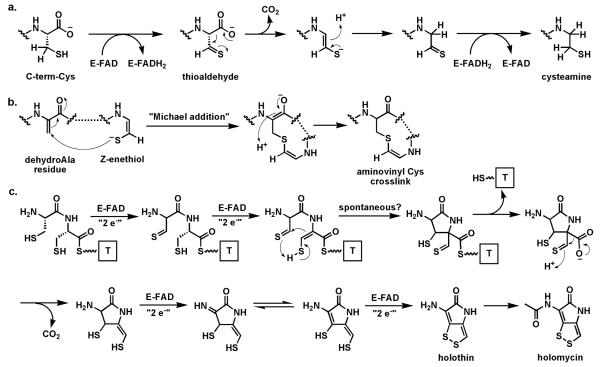
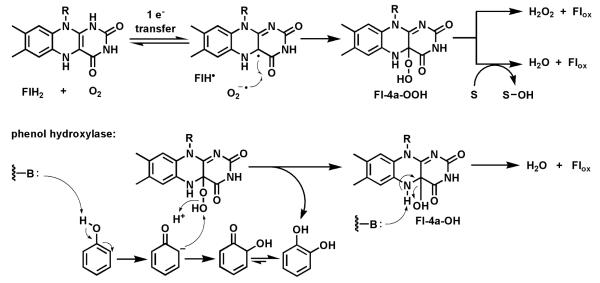
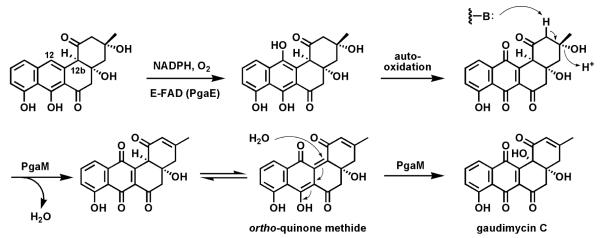
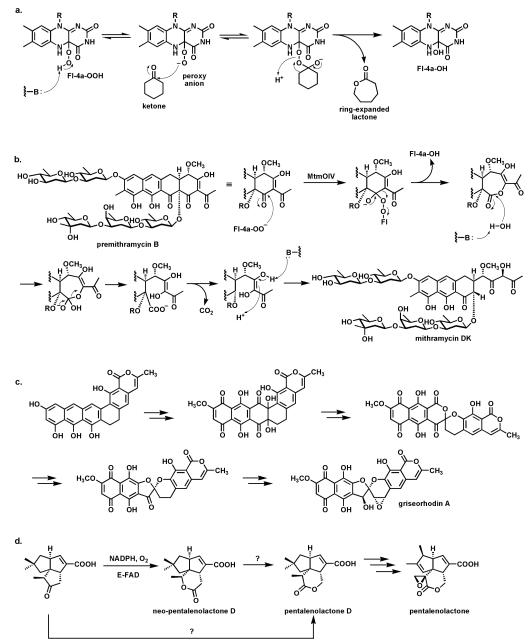
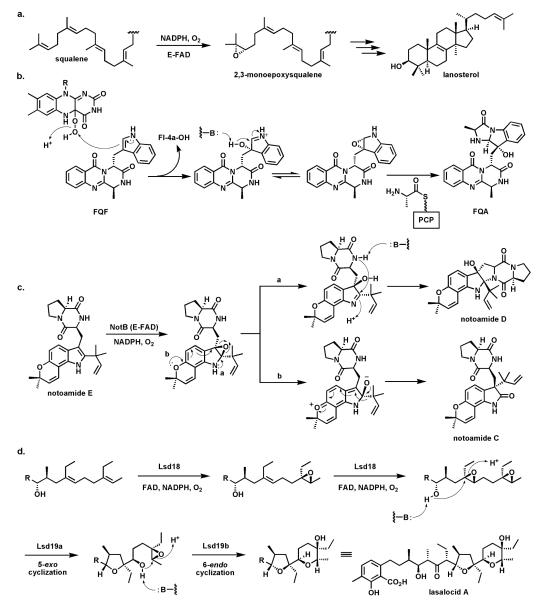
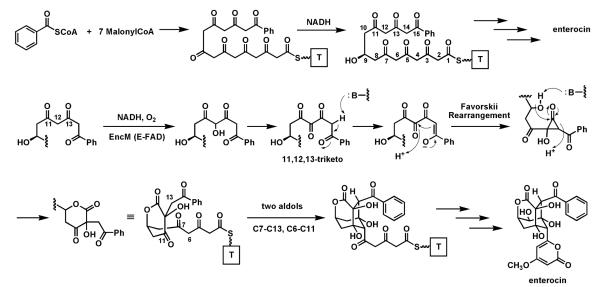
Riboflavin-based coenzymes, tightly bound to enzymes catalyzing substrate oxidations and reductions, enable an enormous range of chemical transformations in biosynthetic pathways. Flavoenzymes catalyze substrate oxidations involving amine and alcohol oxidations and desaturations to olefins, the latter setting up Diels-Alder cyclizations in lovastatin and solanapyrone biosyntheses. Both C(4a) and N(5) of the flavin coenzymes are sites for covalent adduct formation. For example, the reactivity of dihydroflavins with molecular oxygen leads to flavin-4a-OOH adducts which then carry out a diverse range of oxygen transfers, including Baeyer-Villiger type ring expansions, olefin epoxidations, halogenations via transient HOCl generation, and an oxidative Favorskii rerrangement during enterocin assembly.
Figures





















References
-
- Fischbach MA, Walsh CT. Chem. Rev. 2006;106:3468–3496. - PubMed
-
- Hertweck C. Angew. Chem. Int. Ed. 2009;48:4688–4716. - PubMed
-
- Sattely ES, Fischbach MA, Walsh CT. Nat. Prod. Rep. 2008;25:757–793. - PubMed
-
- Walsh CT. Acc. Chem. Res. 2008;41:4–10. - PubMed
-
- Kopp F, Mahlert C, Grünewald J, Marahiel MA. J. Am. Chem. Soc. 2006;128:16478–16479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous