

Synthetic zinc finger repressors reduce mutant huntingtin expression in the brain of R6/2 mice
- PMID: 23054839
- PMCID: PMC3494930
- DOI: 10.1073/pnas.1206506109
Synthetic zinc finger repressors reduce mutant huntingtin expression in the brain of R6/2 mice
Abstract
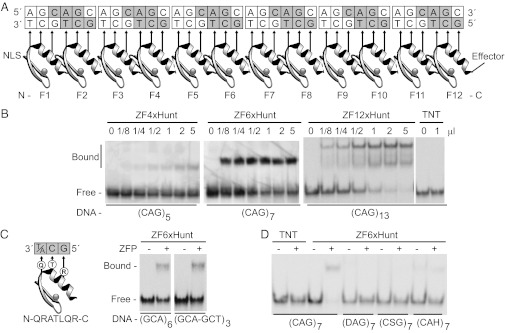
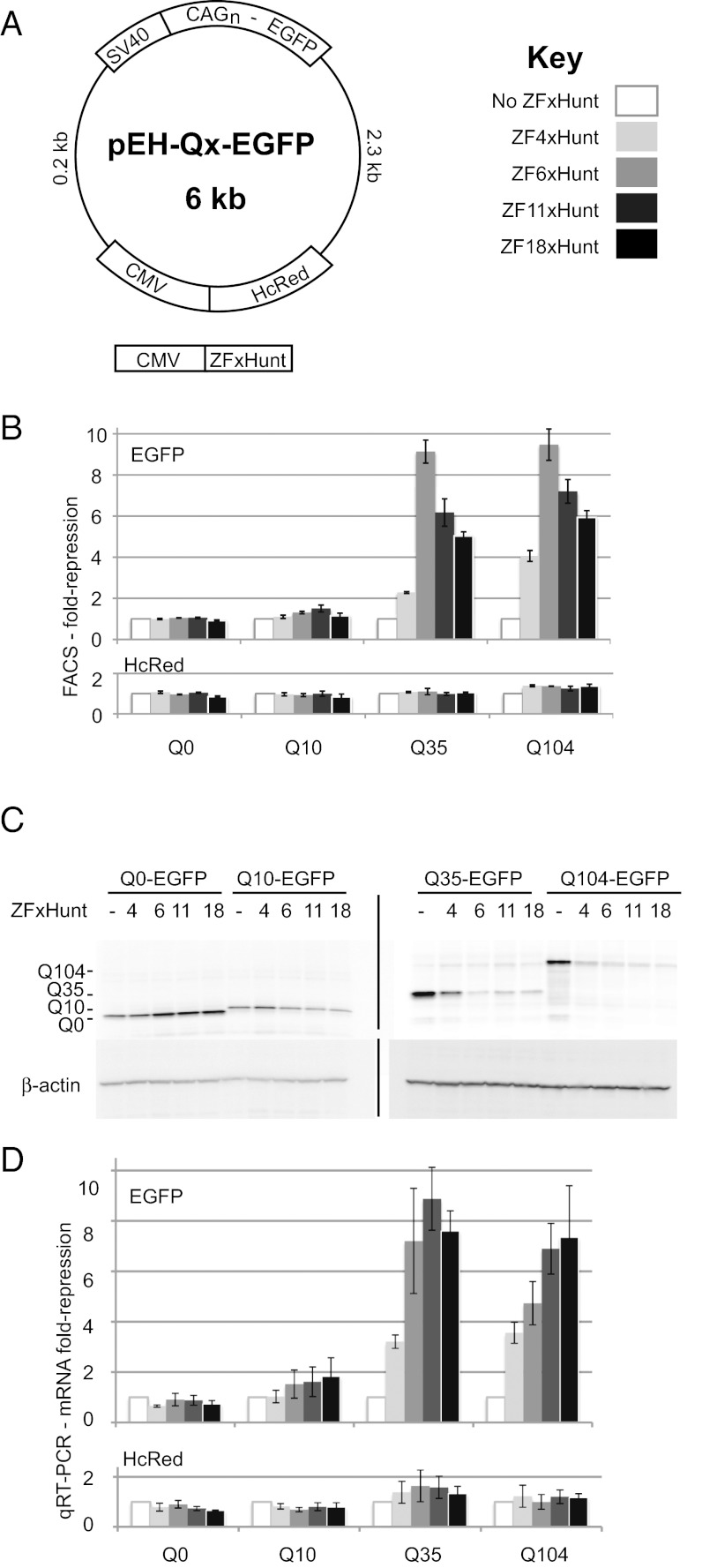
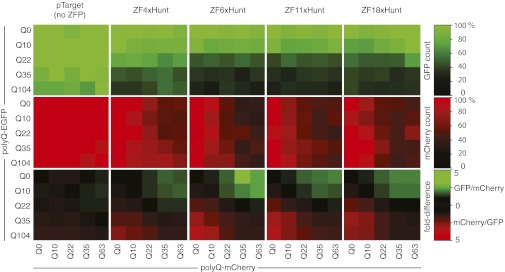
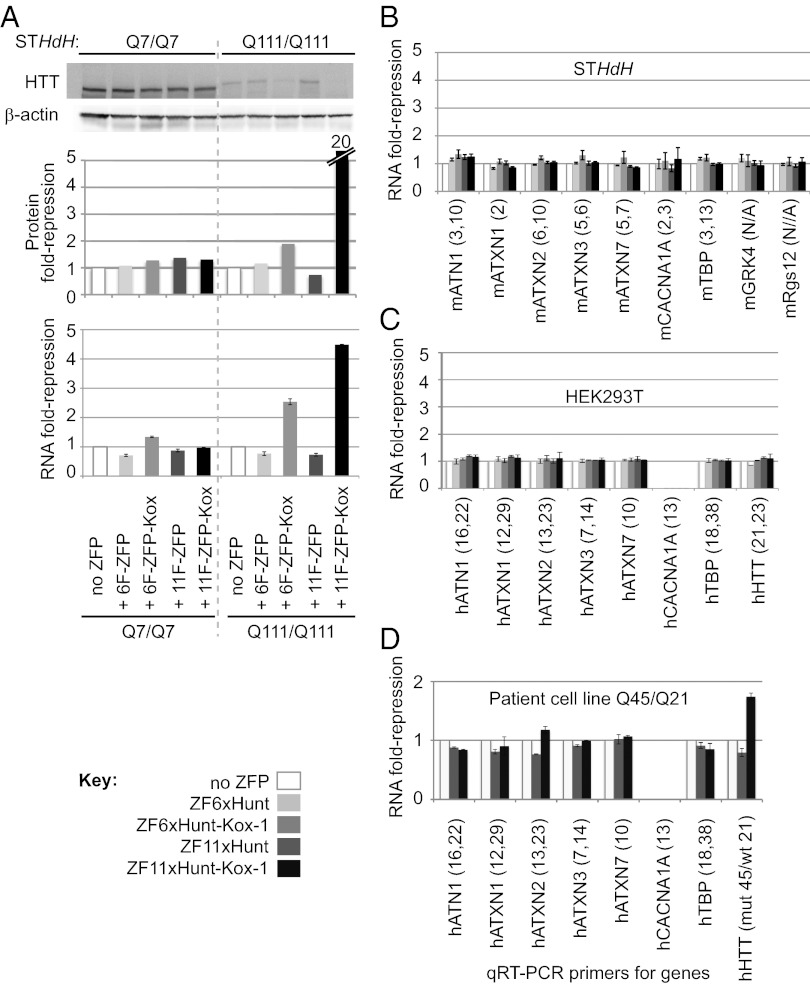
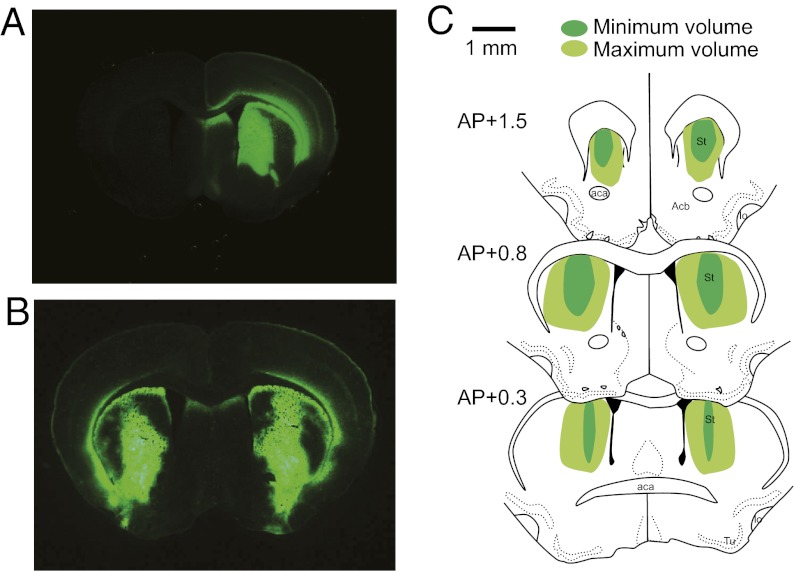
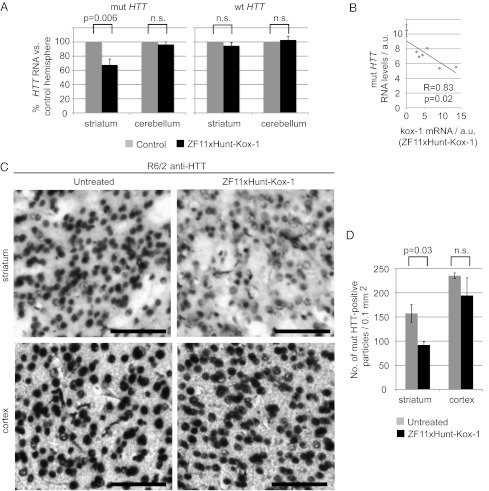
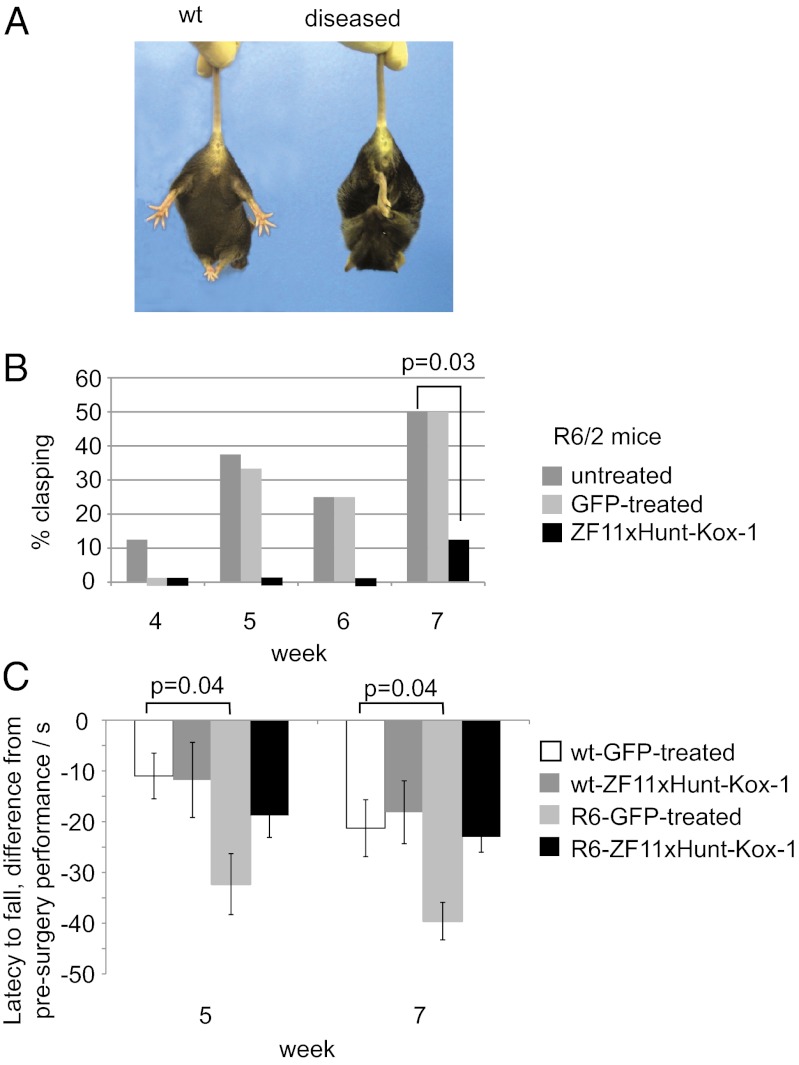
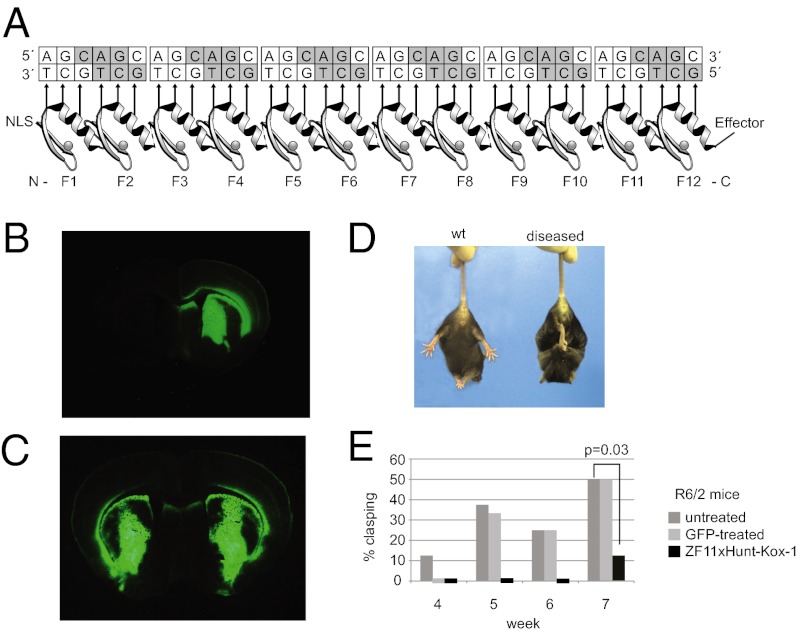
Huntington's disease (HD) is a dominantly inherited neurodegenerative disorder caused by expanded CAG repeats in the huntingtin (HTT) gene. Although several palliative treatments are available, there is currently no cure and patients generally die 10-15 y after diagnosis. Several promising approaches for HD therapy are currently in development, including RNAi and antisense analogs. We developed a complementary strategy to test repression of mutant HTT with zinc finger proteins (ZFPs) in an HD model. We tested a "molecular tape measure" approach, using long artificial ZFP chains, designed to bind longer CAG repeats more strongly than shorter repeats. After optimization, stable ZFP expression in a model HD cell line reduced chromosomal expression of the mutant gene at both the protein and mRNA levels (95% and 78% reduction, respectively). This was achieved chromosomally in the context of endogenous mouse HTT genes, with variable CAG-repeat lengths. Shorter wild-type alleles, other genomic CAG-repeat genes, and neighboring genes were unaffected. In vivo, striatal adeno-associated virus viral delivery in R6/2 mice was efficient and revealed dose-dependent repression of mutant HTT in the brain (up to 60%). Furthermore, zinc finger repression was tested at several levels, resulting in protein aggregate reduction, reduced decline in rotarod performance, and alleviation of clasping in R6/2 mice, establishing a proof-of-principle for synthetic transcription factor repressors in the brain.
Conflict of interest statement
Conflict of interest statement: M.I. and M.G.-C. are co-authors on a patent associated with this technology.
Figures
References
-
- Walker FO. Huntington’s Disease. Semin Neurol. 2007;27(2):143–150. - PubMed
-
- Orr HT, Zoghbi HY. Trinucleotide repeat disorders. Annu Rev Neurosci. 2007;30:575–621. - PubMed
-
- Kumar P, Kalonia H, Kumar A. Huntington’s disease: Pathogenesis to animal models. Pharmacol Rep. 2010;62(1):1–14. - PubMed
-
- The Huntington’s Disease Collaborative Research Group A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell. 1993;72(6):971–983. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
