

Encoding of aversion by dopamine and the nucleus accumbens
- PMID: 23055953
- PMCID: PMC3457027
- DOI: 10.3389/fnins.2012.00137
Encoding of aversion by dopamine and the nucleus accumbens
Abstract
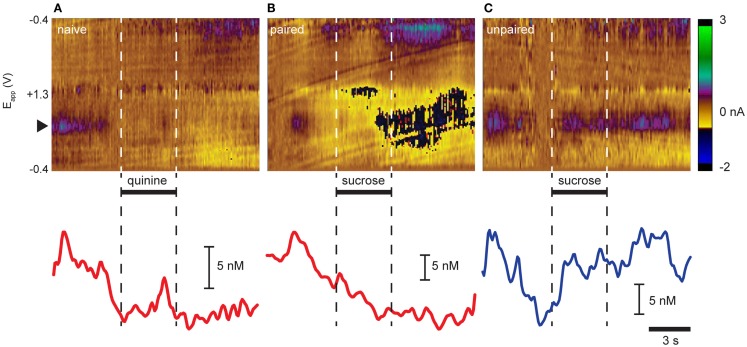
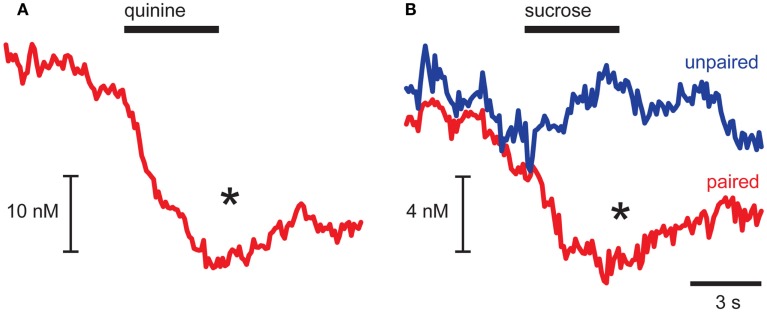
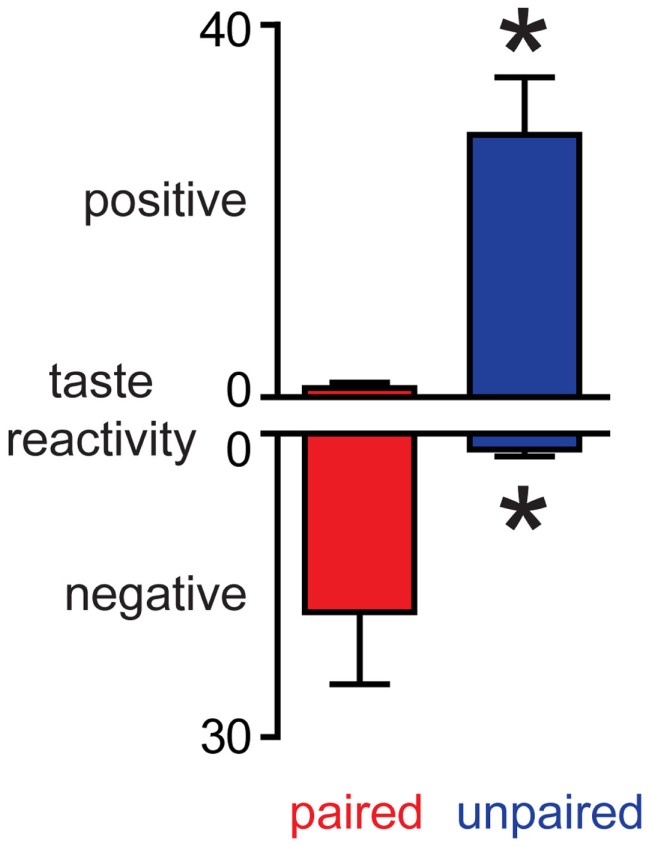
Adaptive motivated behavior requires rapid discrimination between beneficial and harmful stimuli. Such discrimination leads to the generation of either an approach or rejection response, as appropriate, and enables organisms to maximize reward and minimize punishment. Classically, the nucleus accumbens (NAc) and the dopamine projection to it are considered an integral part of the brain's reward circuit, i.e., they direct approach and consumption behaviors and underlie positive reinforcement. This reward-centered framing ignores important evidence about the role of this system in encoding aversive events. One reason for bias toward reward is the difficulty in designing experiments in which animals repeatedly experience punishments; another is the challenge in dissociating the response to an aversive stimulus itself from the reward/relief experienced when an aversive stimulus is terminated. Here, we review studies that employ techniques with sufficient time resolution to measure responses in ventral tegmental area and NAc to aversive stimuli as they are delivered. We also present novel findings showing that the same stimulus - intra-oral infusion of sucrose - has differing effects on NAc shell dopamine release depending on the prior experience. Here, for some rats, sucrose was rendered aversive by explicitly pairing it with malaise in a conditioned taste aversion paradigm. Thereafter, sucrose infusions led to a suppression of dopamine with a similar magnitude and time course to intra-oral infusions of a bitter quinine solution. The results are discussed in the context of regional differences in dopamine signaling and the implications of a pause in phasic dopamine release within the NAc shell. Together with our data, the emerging literature suggests an important role for differential phasic dopamine signaling in aversion vs. reward.
Keywords: conditioned taste aversion; electrophysiology; reward; taste reactivity; ventral tegmental area; voltammetry.
Figures
References
-
- Aragona B. J., Day J. J., Roitman M. F., Cleaveland N. A., Wightman R. M., Carelli R. M. (2009). Regional specificity in the real-time development of phasic dopamine transmission patterns during acquisition of a cue-cocaine association in rats. Eur. J. Neurosci. 30, 1889–1899 10.1111/j.1460-9568.2009.07027.x - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources