

Intron definition and a branch site adenosine at nt 385 control RNA splicing of HPV16 E6*I and E7 expression
- PMID: 23056301
- PMCID: PMC3464268
- DOI: 10.1371/journal.pone.0046412
Intron definition and a branch site adenosine at nt 385 control RNA splicing of HPV16 E6*I and E7 expression
Abstract
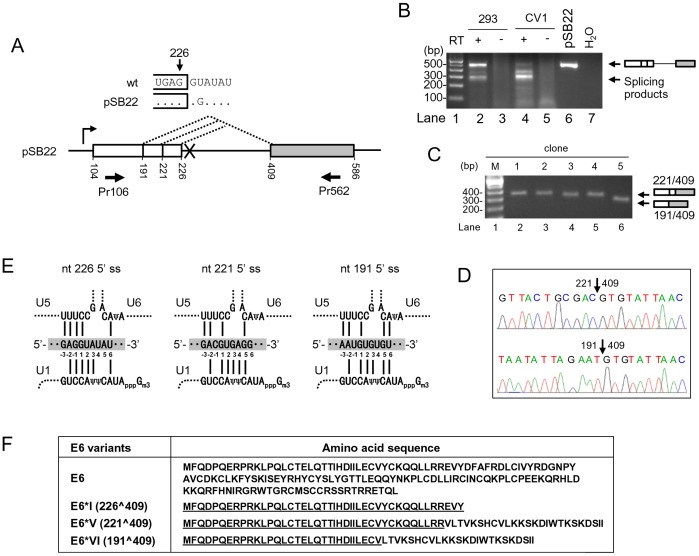
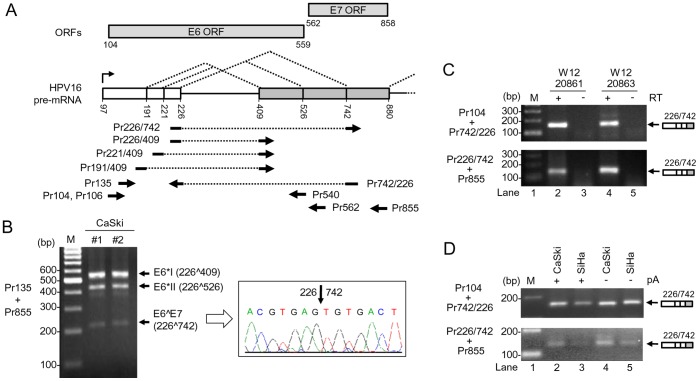
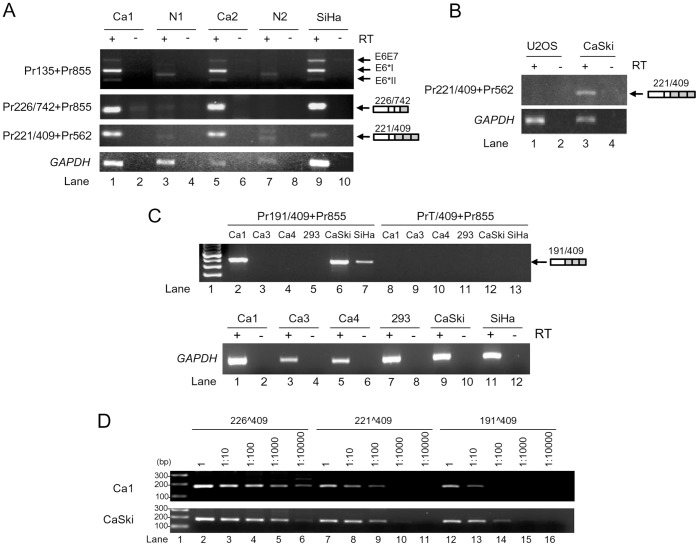
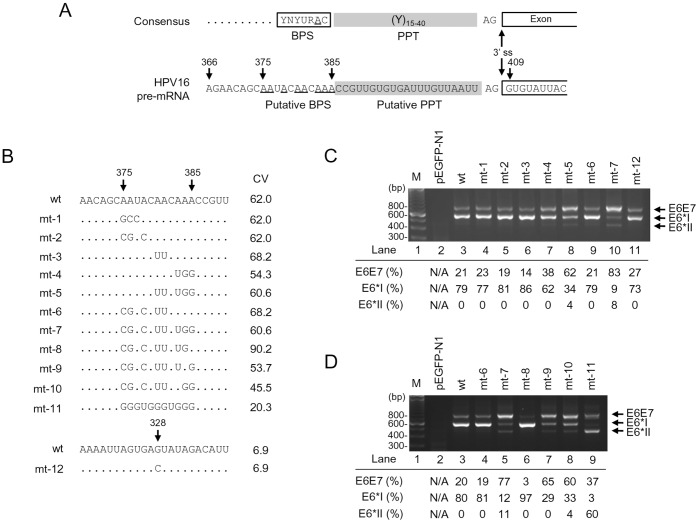
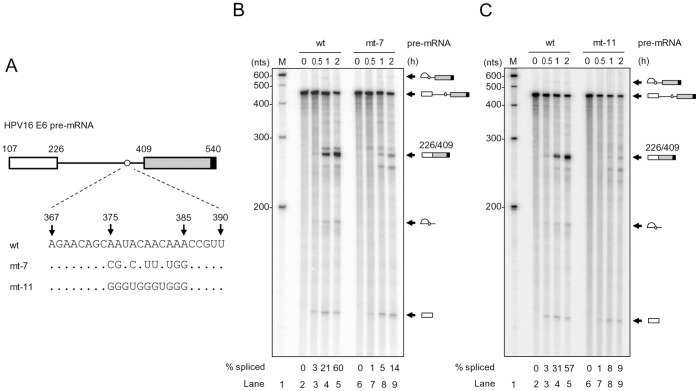
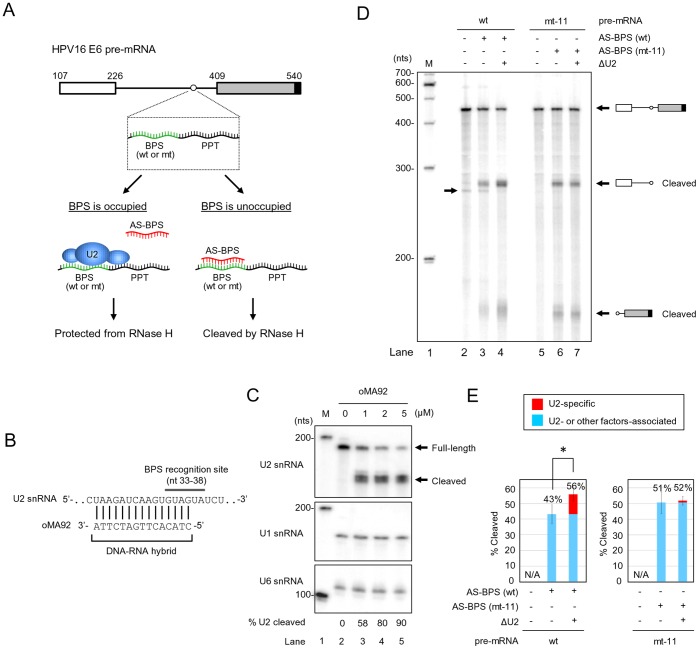
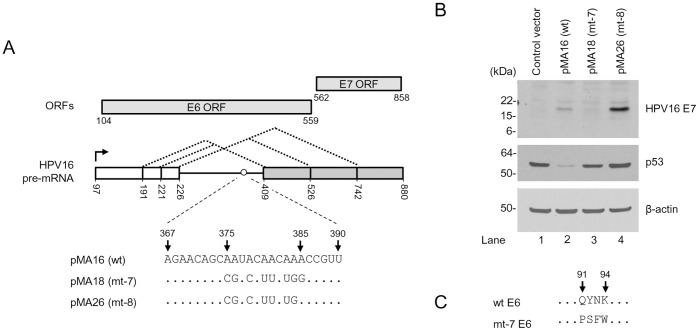
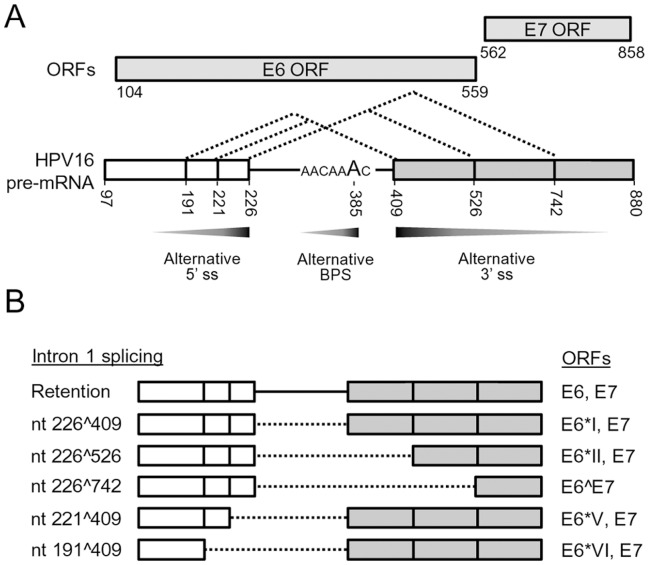
HPV16 E6 and E7, two viral oncogenes, are expressed from a single bicistronic pre-mRNA. In this report, we provide the evidence that the bicistronic pre-mRNA intron 1 contains three 5' splice sites (5' ss) and three 3' splice sites (3' ss) normally used in HPV16(+) cervical cancer and its derived cell lines. The choice of two novel alternative 5' ss (nt 221 5' ss and nt 191 5' ss) produces two novel isoforms of E6E7 mRNAs (E6*V and E6*VI). The nt 226 5' ss and nt 409 3' ss is preferentially selected over the other splice sites crossing over the intron to excise a minimal length of the intron in RNA splicing. We identified AACAAAC as the preferred branch point sequence (BPS) and an adenosine at nt 385 (underlined) in the BPS as a branch site to dictate the selection of the nt 409 3' ss for E6*I splicing and E7 expression. Introduction of point mutations into the mapped BPS led to reduced U2 binding to the BPS and thereby inhibition of the second step of E6E7 splicing at the nt 409 3' ss. Importantly, the E6E7 bicistronic RNA with a mutant BPS and inefficient splicing makes little or no E7 and the resulted E6 with mutations of (91)QYNK(94) to (91)PSFW(94) displays attenuate activity on p53 degradation. Together, our data provide structural basis of the E6E7 intron 1 for better understanding of how viral E6 and E7 expression is regulated by alternative RNA splicing. This study elucidates for the first time a mapped branch point in HPV16 genome involved in viral oncogene expression.
Conflict of interest statement
Figures
References
-
- Howley PM, Lowy DR (2007) Papillomaviruses. In: Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE, eds. Fields Virology. Fifth edition ed. Philadelphia: Lippincott Williams & Wilkins; 2299–2354.
-
- Walboomers JM, Jacobs MV, Manos MM, Bosch FX, Kummer JA, et al. (1999) Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J Pathol 189: 12–19. - PubMed
-
- Munoz N, Bosch FX, de Sanjose S, Herrero R, Castellsague X, et al. (2003) Epidemiologic classification of human papillomavirus types associated with cervical cancer. N Engl J Med 348: 518–527. - PubMed
-
- Munoz N, Castellsague X, de Gonzalez AB, Gissmann L (2006) Chapter 1: HPV in the etiology of human cancer. Vaccine 24 Suppl 3: S3–1–S310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
