

Solution structure, copper binding and backbone dynamics of recombinant Ber e 1-the major allergen from Brazil nut
- PMID: 23056307
- PMCID: PMC3464261
- DOI: 10.1371/journal.pone.0046435
Solution structure, copper binding and backbone dynamics of recombinant Ber e 1-the major allergen from Brazil nut
Abstract
Background: The 2S albumin Ber e 1 is the major allergen in Brazil nuts. Previous findings indicated that the protein alone does not cause an allergenic response in mice, but the addition of components from a Brazil nut lipid fraction were required. Structural details of Ber e 1 may contribute to the understanding of the allergenic properties of the protein and its potential interaction partners.
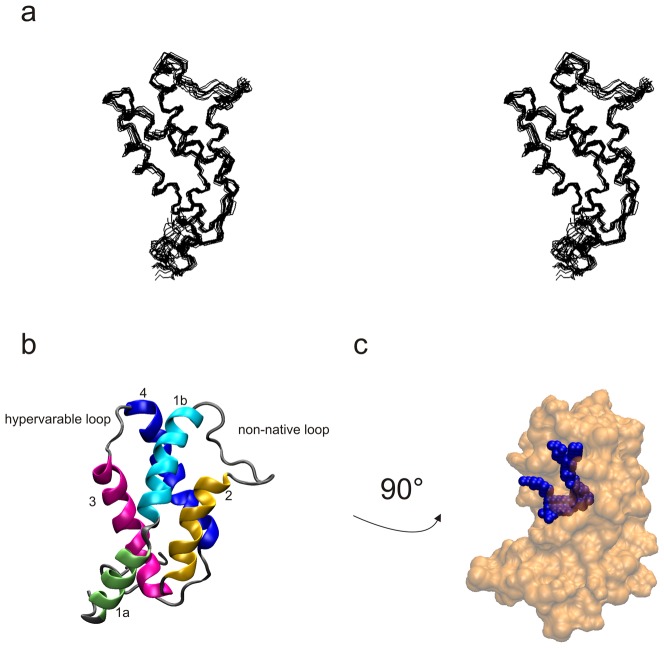
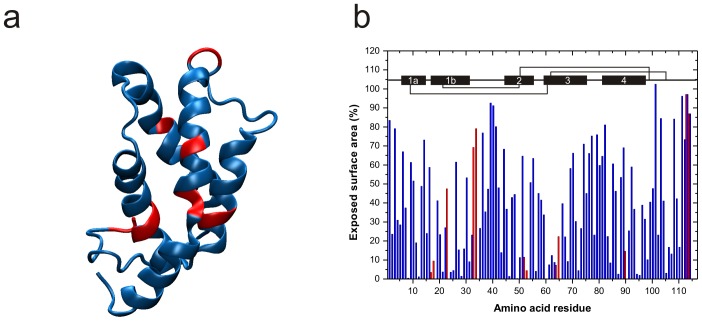
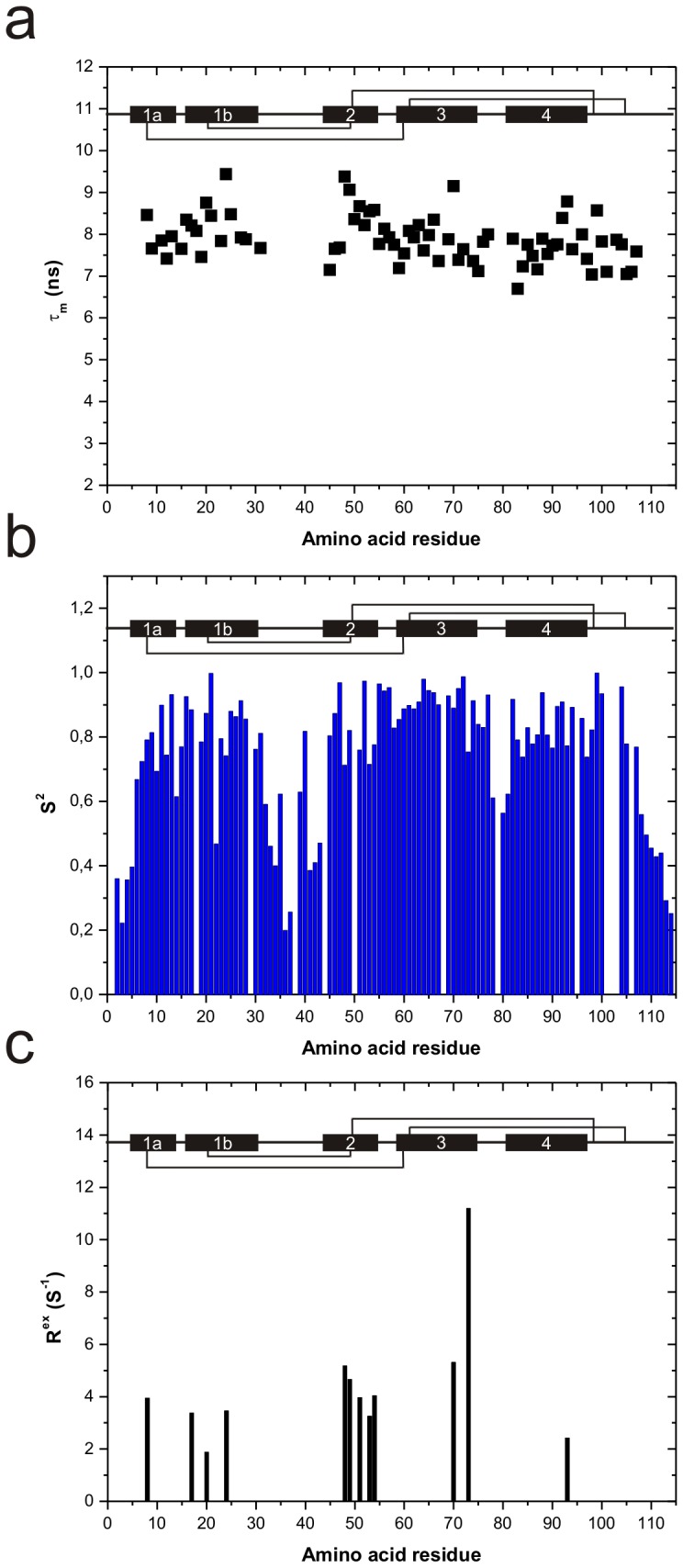
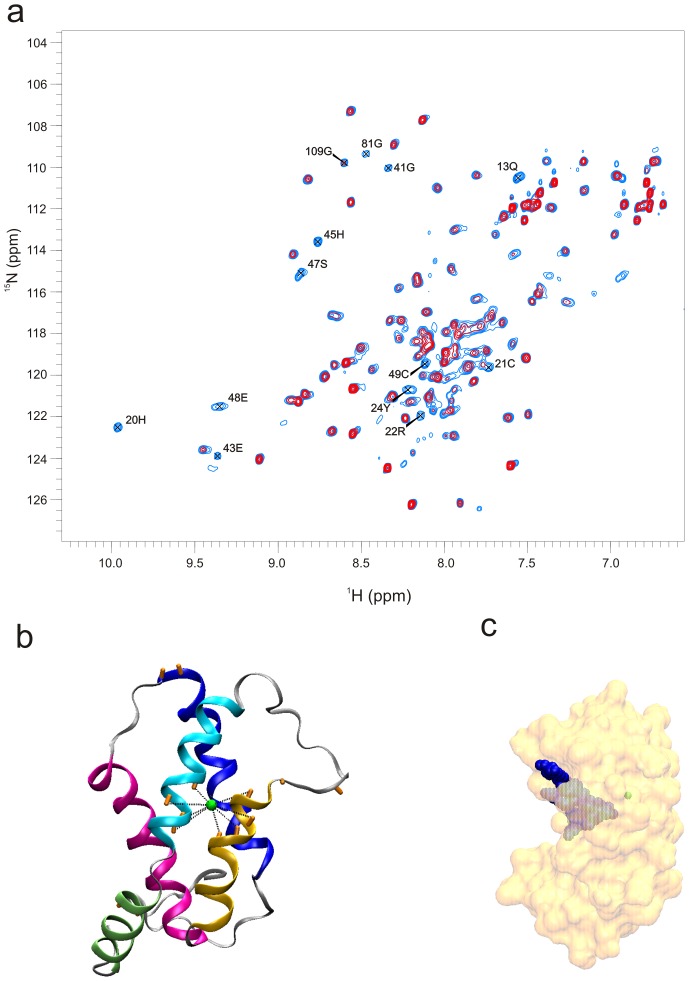
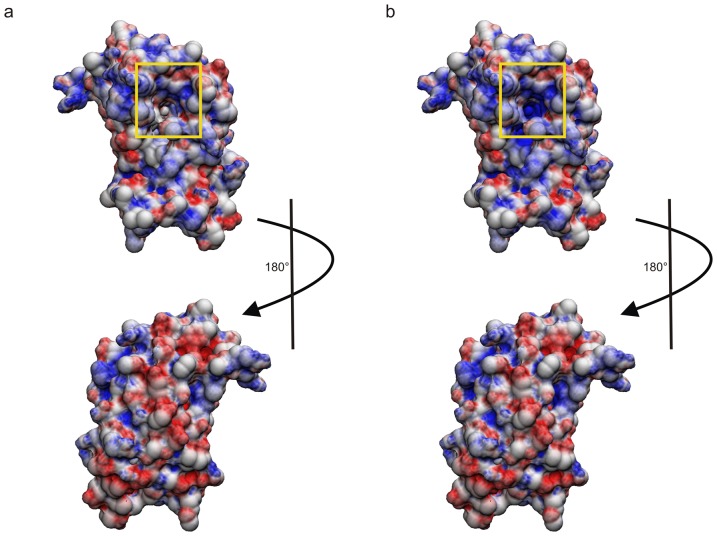
Methodology/principal findings: The solution structure of recombinant Ber e 1 was solved using NMR spectroscopy and measurements of the protein back bone dynamics at a residue-specific level were extracted using (15)N-spin relaxation. A hydrophobic cavity was identified in the structure of Ber e 1. Using the paramagnetic relaxation enhancement property of Cu(2+) in conjunction with NMR, it was shown that Ber e 1 is able to specifically interact with the divalent copper ion and the binding site was modeled into the structure. The IgE binding region as well as the copper binding site show increased dynamics on both fast ps-ns timescale as well as slower µs-ms timescale.
Conclusions/significance: The overall fold of Ber e 1 is similar to other 2S albumins, but the hydrophobic cavity resembles that of a homologous non-specific lipid transfer protein. Ber e 1 is the first 2S albumin shown to interact with Cu(2+) ions. This Cu(2+) binding has minimal effect on the electrostatic potential on the surface of the protein, but the charge distribution within the hydrophobic cavity is significantly altered. As the hydrophobic cavity is likely to be involved in a putative lipid interaction the Cu(2+) can in turn affect the interaction that is essential to provoke an allergenic response.
Conflict of interest statement
Figures
References
-
- Sampson HA (2004) Update on food allergy. J Allergy Clin Immunol 113: 805–819. - PubMed
-
- Radauer C, Bublin M, Wagner S, Mari A, Breiteneder H (2008) Allergens are distributed into few protein families and possess a restricted number of biochemical functions. J Allergy Clin Immunol 121: 847–852 e847. - PubMed
-
- Ruiter B, Shreffler WG (2012) The role of dendritic cells in food allergy. Journal of Allergy and Clinical Immunology 129: 921–928. - PubMed
-
- Radauer C, Breiteneder H (2007) Evolutionary biology of plant food allergens. J Allergy Clin Immunol 120: 518–525. - PubMed
-
- Kreis M, Forde BG, Rahman S, Miflin BJ, Shewry PR (1985) Molecular evolution of the seed storage proteins of barley, rye and wheat. J Mol Biol 183: 499–502. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
