

The genome of Chelonid herpesvirus 5 harbors atypical genes
- PMID: 23056373
- PMCID: PMC3462797
- DOI: 10.1371/journal.pone.0046623
The genome of Chelonid herpesvirus 5 harbors atypical genes
Abstract
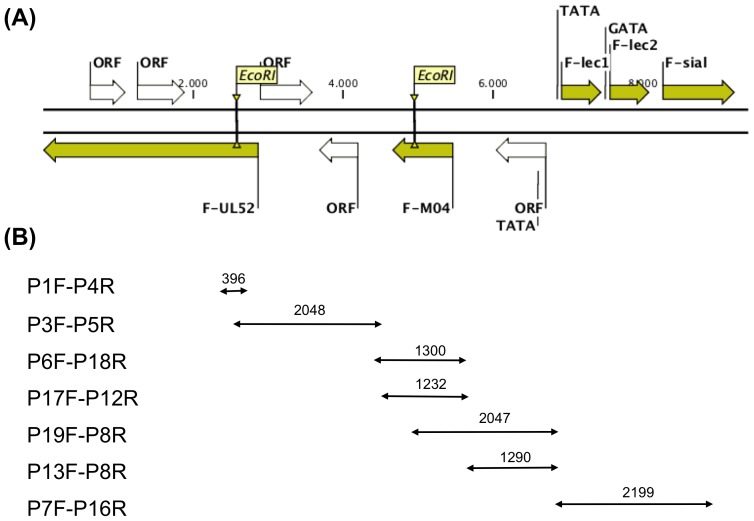
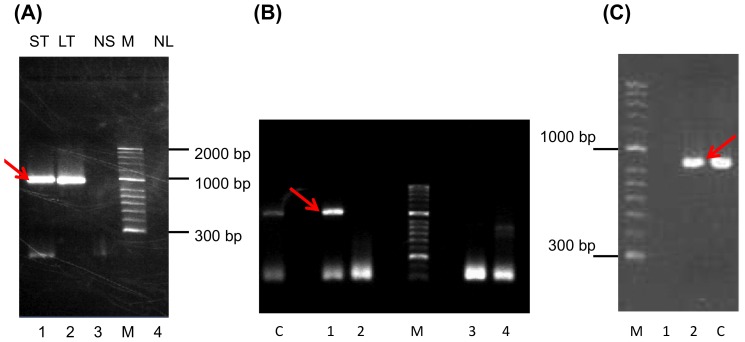
The Chelonid fibropapilloma-associated herpesvirus (CFPHV; ChHV5) is believed to be the causative agent of fibropapillomatosis (FP), a neoplastic disease of marine turtles. While clinical signs and pathology of FP are well known, research on ChHV5 has been impeded because no cell culture system for its propagation exists. We have cloned a BAC containing ChHV5 in pTARBAC2.1 and determined its nucleotide sequence. Accordingly, ChHV5 has a type D genome and its predominant gene order is typical for the varicellovirus genus within the alphaherpesvirinae. However, at least four genes that are atypical for an alphaherpesvirus genome were also detected, i.e. two members of the C-type lectin-like domain superfamily (F-lec1, F-lec2), an orthologue to the mouse cytomegalovirus M04 (F-M04) and a viral sialyltransferase (F-sial). Four lines of evidence suggest that these atypical genes are truly part of the ChHV5 genome: (1) the pTARBAC insertion interrupted the UL52 ORF, leaving parts of the gene to either side of the insertion and suggesting that an intact molecule had been cloned. (2) Using FP-associated UL52 (F-UL52) as an anchor and the BAC-derived sequences as a means to generate primers, overlapping PCR was performed with tumor-derived DNA as template, which confirmed the presence of the same stretch of "atypical" DNA in independent FP cases. (3) Pyrosequencing of DNA from independent tumors did not reveal previously undetected viral sequences, suggesting that no apparent loss of viral sequence had happened due to the cloning strategy. (4) The simultaneous presence of previously known ChHV5 sequences and F-sial as well as F-M04 sequences was also confirmed in geographically distinct Australian cases of FP. Finally, transcripts of F-sial and F-M04 but not transcripts of lytic viral genes were detected in tumors from Hawaiian FP-cases. Therefore, we suggest that F-sial and F-M04 may play a role in FP pathogenesis.
Conflict of interest statement
Figures


References
-
- Herbst L, Ene A, Su M, Desalle R, Lenz J (2004) Tumor outbreaks in marine turtles are not due to recent herpesvirus mutations. Curr Biol 14: R697–699. - PubMed
-
- Quackenbush SL, Work TM, Balazs GH, Casey RN, Rovnak J, et al. (1998) Three closely related herpesviruses are associated with fibropapillomatosis in marine turtles. Virology 246: 392–399. - PubMed
-
- Smith GM, Coates CW (1938) Fibro-epithelial growths of the skin in large marine turtles, Chelonia mydas (Linneaus). Zoologica (NY) 23: 93–98.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
