

Cardiac regenerative capacity and mechanisms
- PMID: 23057748
- PMCID: PMC3586268
- DOI: 10.1146/annurev-cellbio-101011-155739
Cardiac regenerative capacity and mechanisms
Abstract
The heart holds the monumental yet monotonous task of maintaining circulation. Although cardiac function is critical to other organs and to life itself, mammals are not equipped with significant natural capacity to replace heart muscle that has been lost by injury. This deficiency plays a role in leaving millions worldwide vulnerable to heart failure each year. By contrast, certain other vertebrate species such as zebrafish are strikingly good at heart regeneration. A cellular and molecular understanding of endogenous regenerative mechanisms and advances in methodology to transplant cells together project a future in which cardiac muscle regeneration can be therapeutically stimulated in injured human hearts. This review focuses on what has been discovered recently about cardiac regenerative capacity and how natural mechanisms of heart regeneration in model systems are stimulated and maintained.
Figures
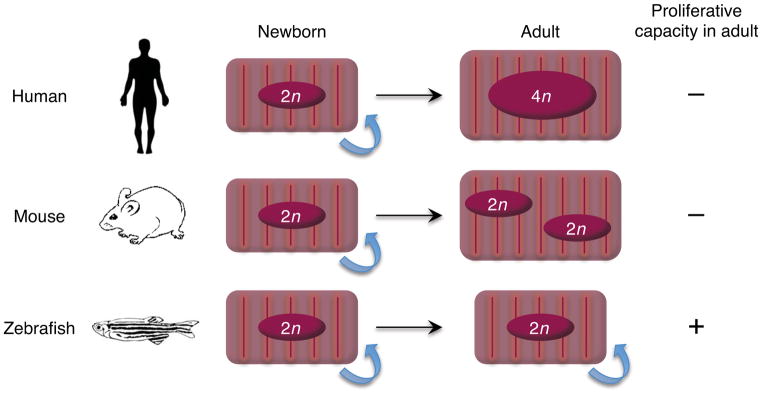
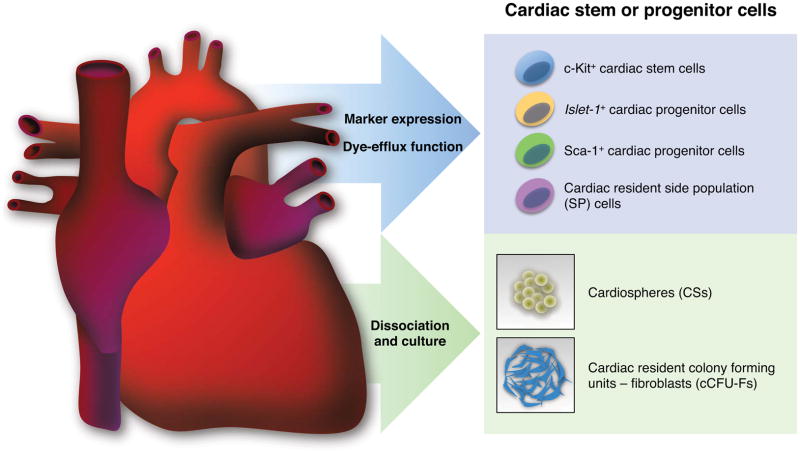
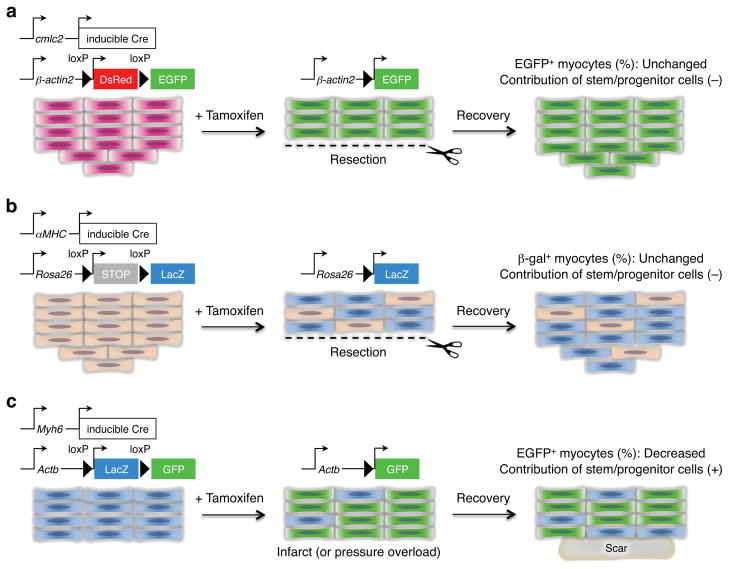
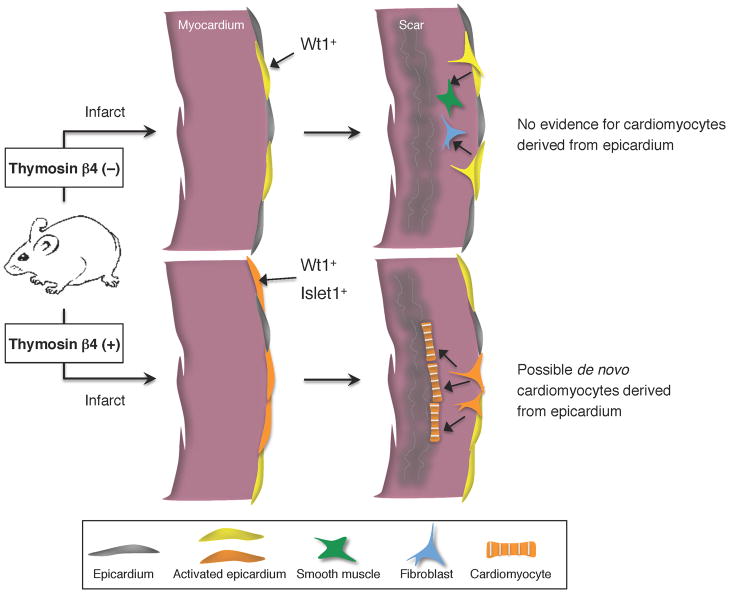
References
-
- Bader D, Oberpriller JO. Repair and reorganization of minced cardiac muscle in the adult newt (Notophthalmus viridescens) Journal of Morphology. 1978;155:349–57. - PubMed
-
- Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature. 2004;428:668–73. - PubMed
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–07. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
