

The unanticipated complexity of the selectivity-filter glutamates of nicotinic receptors
- PMID: 23064317
- PMCID: PMC3508336
- DOI: 10.1038/nchembio.1092
The unanticipated complexity of the selectivity-filter glutamates of nicotinic receptors
Abstract
In ion channels, 'rings' of ionized side chains that decorate the walls of the permeation pathway often lower the energetic barrier to ion conduction. Using single-channel electrophysiological recordings, we studied the poorly understood ring of four glutamates (and one glutamine) that dominates this catalytic effect in the muscle nicotinic acetylcholine receptor ('the intermediate ring of charge'). We show that all four wild-type glutamate side chains are deprotonated in the range of 6.0-9.0 pH, that only two of them contribute to the size of the single-channel current, that these side chains must be able to adopt alternate conformations that either allow or prevent their negative charges from increasing the rate of cation conduction and that the location of these glutamate side chains squarely at one of the ends of the transmembrane pore is critical for their largely unshifted pK(a) values and for the unanticipated impact of their conformational flexibility on cation permeation.
Conflict of interest statement
Figures

 ,
,
 ,
,
 and
and
 ), i–V data from all possible combinations of one glutamate and four alanines around the ring were recorded, and these are indicated with different symbols (one glutamate in either α1 subunit:
; one in β1:
; one in δ:
; one in ε:
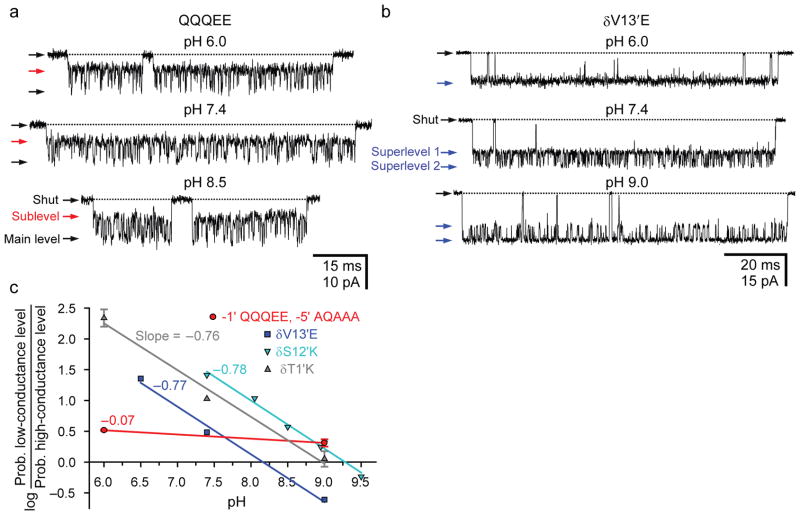
). For the data in this graph, whenever the ε subunit carried a (mutant) glutamate at position –1′, the “extra” glycine occupying position –2′ (or –3′; see Supplementary Fig. 1a) was deleted. b, Single-channel inward currents (outside-out configuration; 1μM ACh; solutions 3 and 4) recorded from the adult muscle AChR carrying the burst-prolongingεT264P mutation. The pH values of the extra- and intracellular solutions were the same. For the traces shown, the applied potential was −110 mV at pH 6.0, −100 mV at pH 7.4 and −80 mV at pH 9.0. Openings are downward deflections. Display fc ≅ 6 kHz.
), i–V data from all possible combinations of one glutamate and four alanines around the ring were recorded, and these are indicated with different symbols (one glutamate in either α1 subunit:
; one in β1:
; one in δ:
; one in ε:
). For the data in this graph, whenever the ε subunit carried a (mutant) glutamate at position –1′, the “extra” glycine occupying position –2′ (or –3′; see Supplementary Fig. 1a) was deleted. b, Single-channel inward currents (outside-out configuration; 1μM ACh; solutions 3 and 4) recorded from the adult muscle AChR carrying the burst-prolongingεT264P mutation. The pH values of the extra- and intracellular solutions were the same. For the traces shown, the applied potential was −110 mV at pH 6.0, −100 mV at pH 7.4 and −80 mV at pH 9.0. Openings are downward deflections. Display fc ≅ 6 kHz.
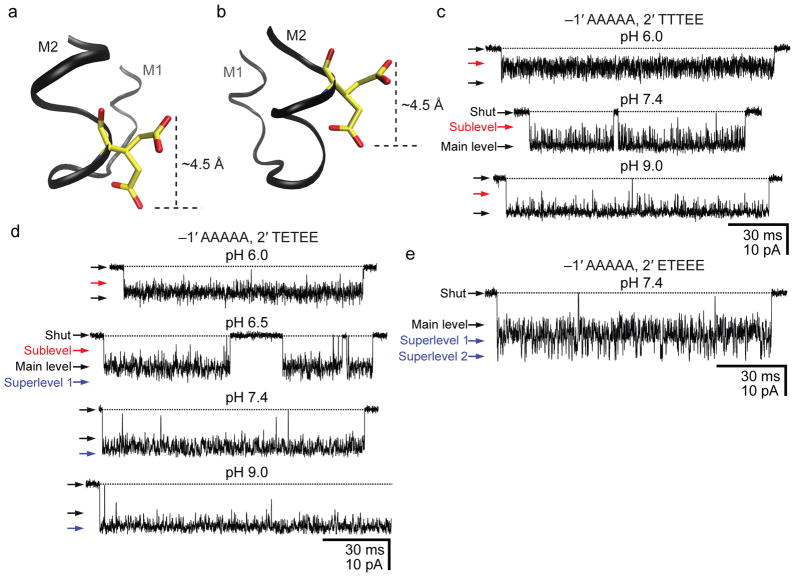
 ; outside-out configuration; −150 mV; 1 μM ACh; symmetrical pH; solutions 3 and 5) and from the channel with a glutamate engineered at position δ13′ (
; outside-out configuration; −150 mV; 1 μM ACh; symmetrical pH; solutions 3 and 5) and from the channel with a glutamate engineered at position δ13′ (
 ; cell-attached configuration; ~−100 mV; 1 μM ACh; solutions 1 and 2). Data corresponding to open-channel current fluctuations recorded from two δM2 lysine mutants (
and
; cell-attached configuration; ~−100 mV; 1 μM ACh; solutions 1 and 2). Data corresponding to open-channel current fluctuations recorded from two δM2 lysine mutants (
and
 ; outside-out configuration; −100 mV; 1 μM ACh; symmetrical pH; solutions 3 and 5) are also included, for comparison.
; outside-out configuration; −100 mV; 1 μM ACh; symmetrical pH; solutions 3 and 5) are also included, for comparison.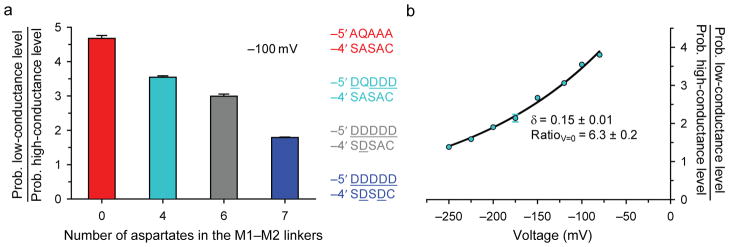
Comment in
-
Channels: Rotamers affect ion conductance.Nat Chem Biol. 2012 Dec;8(12):953-4. doi: 10.1038/nchembio.1115. Nat Chem Biol. 2012. PMID: 23183578 Free PMC article.
References
-
- Imoto K, et al. Rings of negatively charged amino acids determine the acetylcholine receptor channel conductance. Nature. 1988;335:645–648. - PubMed
-
- Galzi JL, et al. Mutations in the ion channel domain of a neuronal nicotinic receptor convert ion selectivity from cationic to anionic. Nature. 1992;359:500–505. - PubMed
-
- Corringer PJ, et al. Mutational analysis of the charge selectivity filter of the α7 nicotinic acetylcholine receptor. Neuron. 1999;22:831–843. - PubMed
-
- Prod’hom B, Pietrobon D, Hess P. Direct measurement of proton transfer rates to a group controlling the dihydropyridine-sensitive Ca2+ channel. Nature. 1987;329:243–246. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
