

DNA polymerase delta in DNA replication and genome maintenance
- PMID: 23065663
- PMCID: PMC3694620
- DOI: 10.1002/em.21745
DNA polymerase delta in DNA replication and genome maintenance
Abstract
The eukaryotic genome is in a constant state of modification and repair. Faithful transmission of the genomic information from parent to daughter cells depends upon an extensive system of surveillance, signaling, and DNA repair, as well as accurate synthesis of DNA during replication. Often, replicative synthesis occurs over regions of DNA that have not yet been repaired, presenting further challenges to genomic stability. DNA polymerase δ (pol δ) occupies a central role in all of these processes: catalyzing the accurate replication of a majority of the genome, participating in several DNA repair synthetic pathways, and contributing structurally to the accurate bypass of problematic lesions during translesion synthesis. The concerted actions of pol δ on the lagging strand, pol ϵ on the leading strand, associated replicative factors, and the mismatch repair (MMR) proteins results in a mutation rate of less than one misincorporation per genome per replication cycle. This low mutation rate provides a high level of protection against genetic defects during development and may prevent the initiation of malignancies in somatic cells. This review explores the role of pol δ in replication fidelity and genome maintenance.
Copyright © 2012 Wiley Periodicals, Inc.
Figures
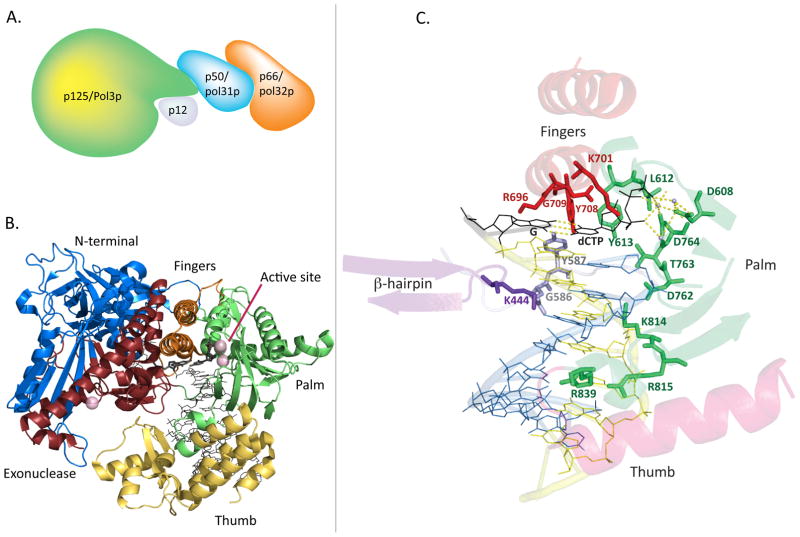
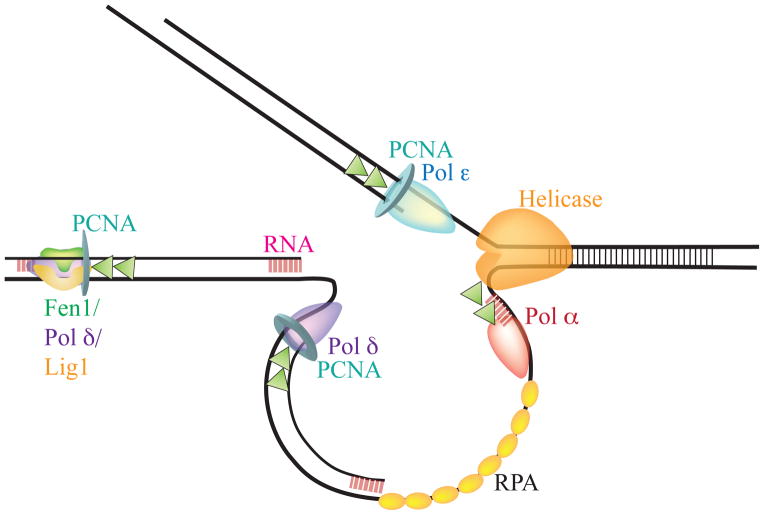
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
