

Tug-of-war in motor protein ensembles revealed with a programmable DNA origami scaffold
- PMID: 23065903
- PMCID: PMC3840815
- DOI: 10.1126/science.1226734
Tug-of-war in motor protein ensembles revealed with a programmable DNA origami scaffold
Abstract
Cytoplasmic dynein and kinesin-1 are microtubule-based motors with opposite polarity that transport a wide variety of cargo in eukaryotic cells. Many cellular cargos demonstrate bidirectional movement due to the presence of ensembles of dynein and kinesin, but are ultimately sorted with spatial and temporal precision. To investigate the mechanisms that coordinate motor ensemble behavior, we built a programmable synthetic cargo using three-dimensional DNA origami to which varying numbers of DNA oligonucleotide-linked motors could be attached, allowing for control of motor type, number, spacing, and orientation in vitro. In ensembles of one to seven identical-polarity motors, motor number had minimal affect on directional velocity, whereas ensembles of opposite-polarity motors engaged in a tug-of-war resolvable by disengaging one motor species.
Figures
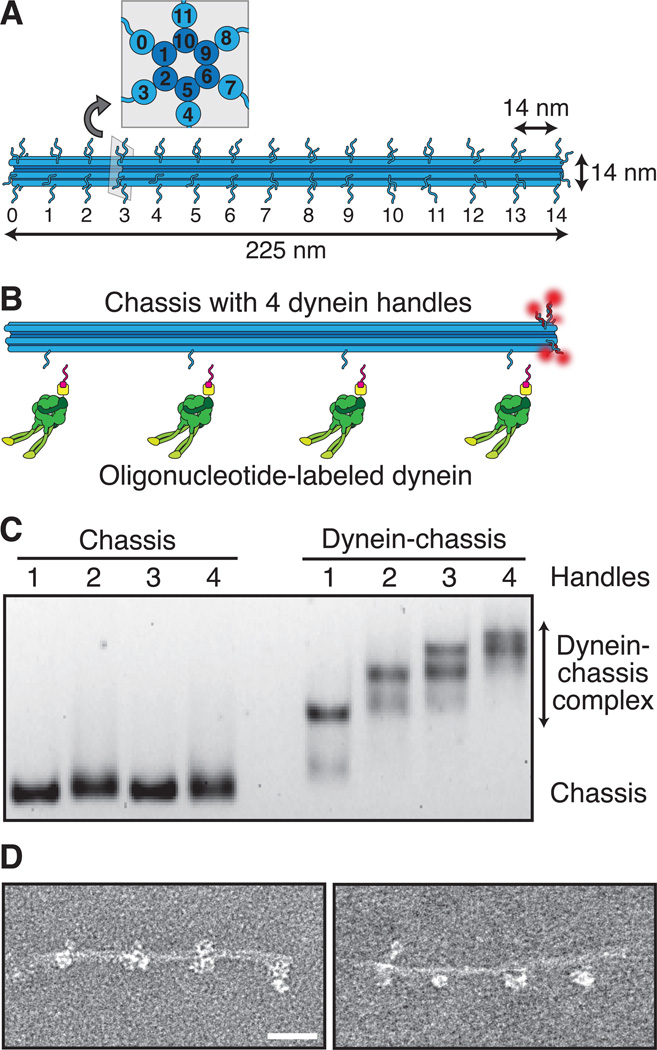
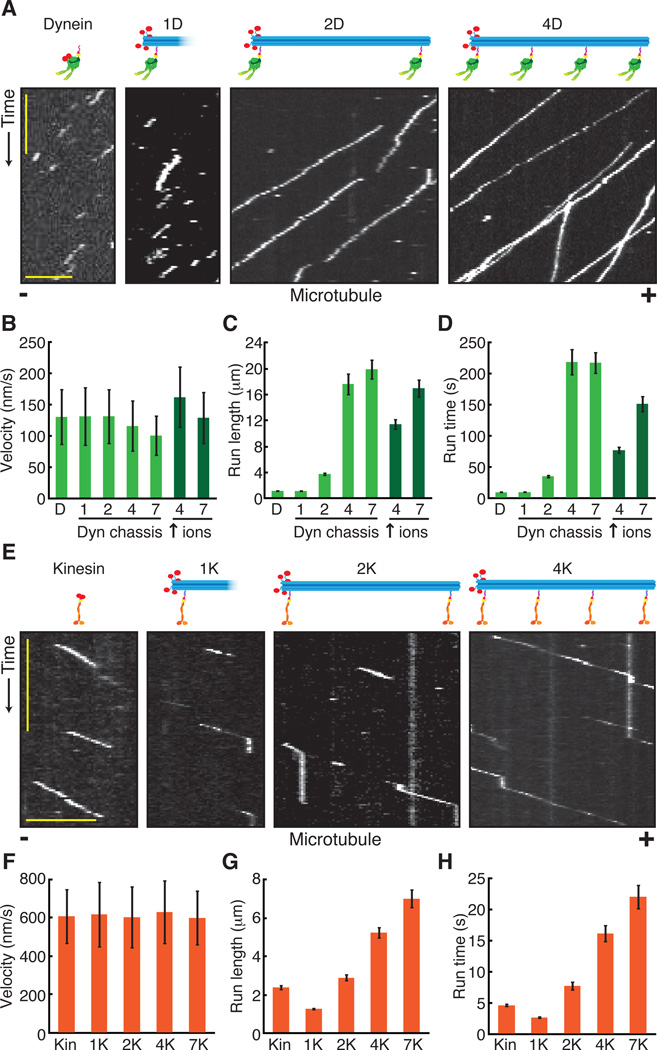
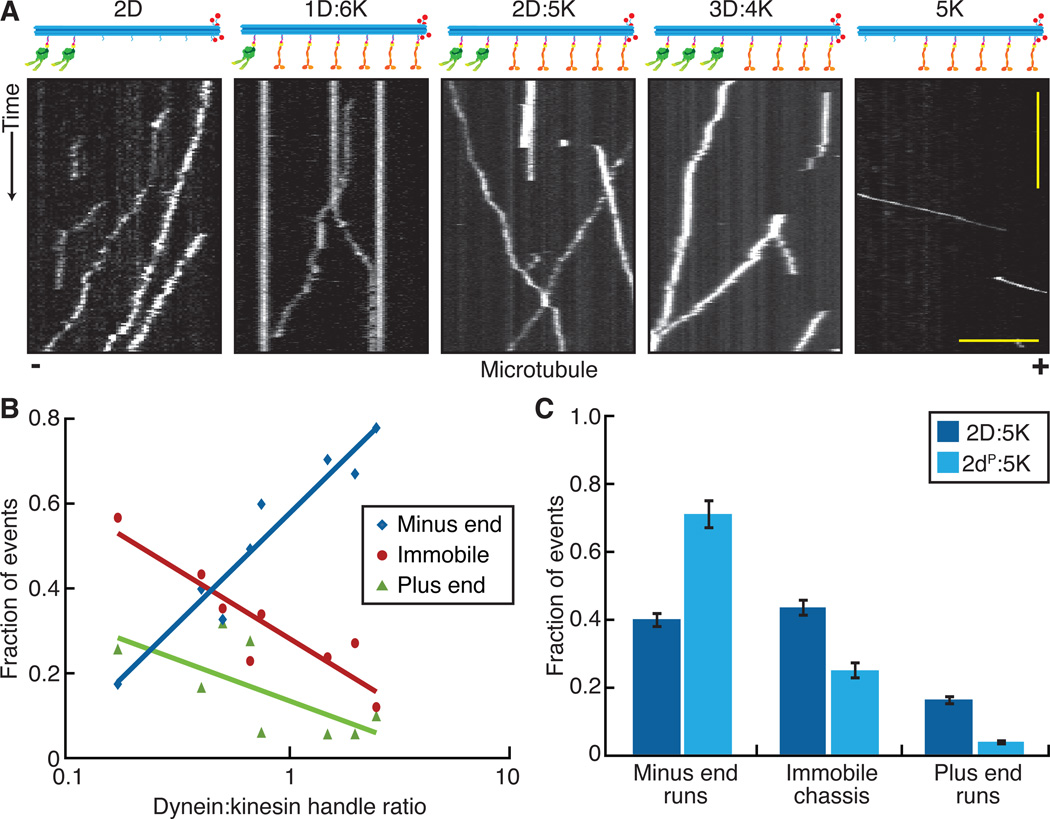
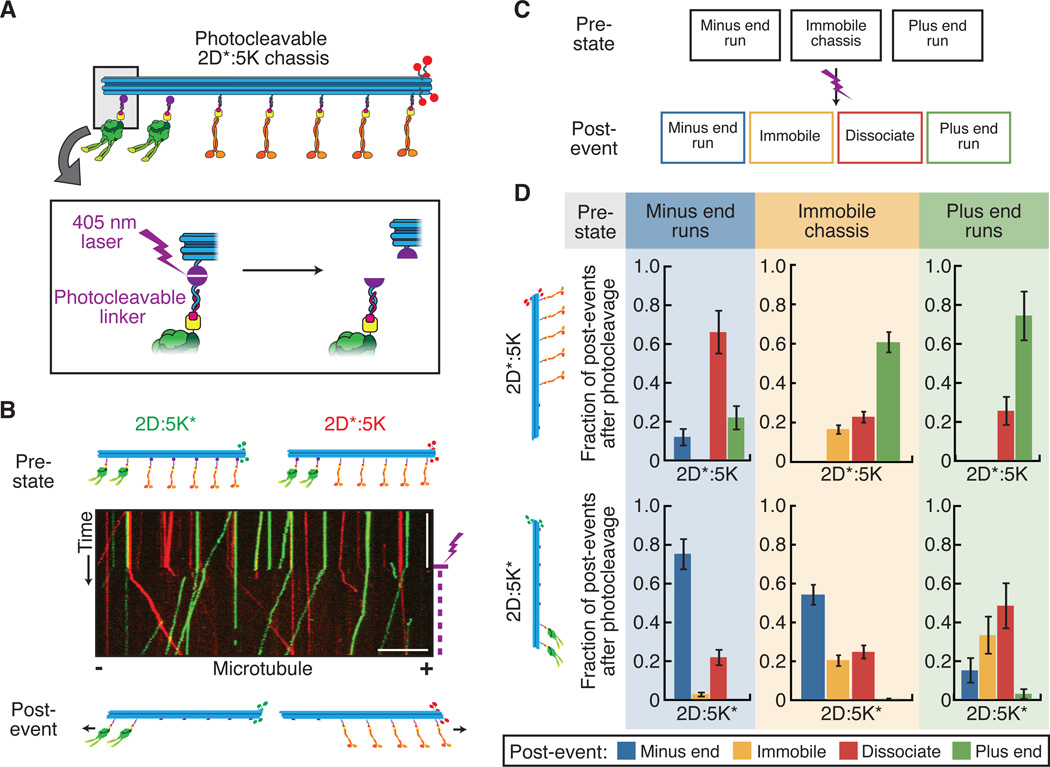
Comment in
-
Biochemistry. Templating a molecular tug-of-war.Science. 2012 Nov 2;338(6107):626-7. doi: 10.1126/science.1230818. Science. 2012. PMID: 23118181 No abstract available.
Similar articles
-
Kinesin-1, -2, and -3 motors use family-specific mechanochemical strategies to effectively compete with dynein during bidirectional transport.Elife. 2022 Sep 20;11:e82228. doi: 10.7554/eLife.82228. Elife. 2022. PMID: 36125250 Free PMC article.
-
Engineering defined motor ensembles with DNA origami.Methods Enzymol. 2014;540:169-88. doi: 10.1016/B978-0-12-397924-7.00010-8. Methods Enzymol. 2014. PMID: 24630107 Review.
-
Cargo rigidity affects the sensitivity of dynein ensembles to individual motor pausing.Cytoskeleton (Hoboken). 2016 Dec;73(12):693-702. doi: 10.1002/cm.21339. Epub 2016 Nov 8. Cytoskeleton (Hoboken). 2016. PMID: 27718534
-
Tug-of-war of microtubule filaments at the boundary of a kinesin- and dynein-patterned surface.Sci Rep. 2014 Jun 13;4:5281. doi: 10.1038/srep05281. Sci Rep. 2014. PMID: 24923426 Free PMC article.
-
Directionality of kinesin motors.Acta Biochim Pol. 2002;49(4):813-21. Acta Biochim Pol. 2002. PMID: 12545188 Review.
Cited by
-
Three-dimensional structures self-assembled from DNA bricks.Science. 2012 Nov 30;338(6111):1177-83. doi: 10.1126/science.1227268. Science. 2012. PMID: 23197527 Free PMC article.
-
Tuning myosin-driven sorting on cellular actin networks.Elife. 2015 Mar 4;4:e05472. doi: 10.7554/eLife.05472. Elife. 2015. PMID: 25738229 Free PMC article.
-
Cryo-Electron Microscopy and Mass Analysis of Oligolysine-Coated DNA Nanostructures.ACS Nano. 2021 Jun 22;15(6):9391-9403. doi: 10.1021/acsnano.0c10137. Epub 2021 Mar 16. ACS Nano. 2021. PMID: 33724780 Free PMC article.
-
Samara Reck-Peterson: dynein defies expectations. Interview by Caitlin Sedwick.J Cell Biol. 2013 Jan 21;200(2):128-9. doi: 10.1083/jcb.2002pi. J Cell Biol. 2013. PMID: 23337114 Free PMC article.
-
JIP1 regulates the directionality of APP axonal transport by coordinating kinesin and dynein motors.J Cell Biol. 2013 Aug 5;202(3):495-508. doi: 10.1083/jcb.201302078. Epub 2013 Jul 29. J Cell Biol. 2013. PMID: 23897889 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
