

Telomere length homeostasis and telomere position effect on a linear human artificial chromosome are dictated by the genetic background
- PMID: 23066103
- PMCID: PMC3526267
- DOI: 10.1093/nar/gks926
Telomere length homeostasis and telomere position effect on a linear human artificial chromosome are dictated by the genetic background
Abstract
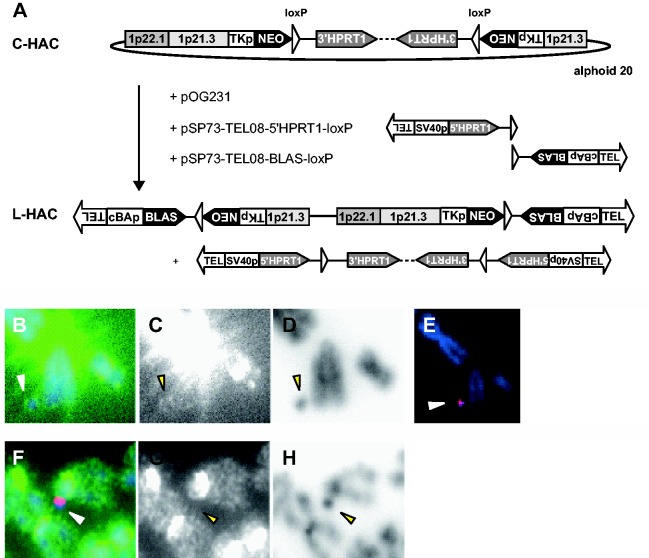
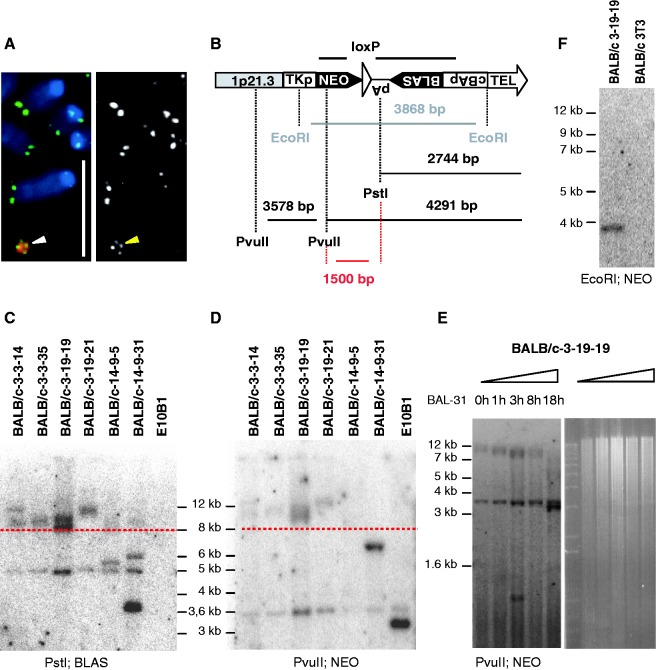
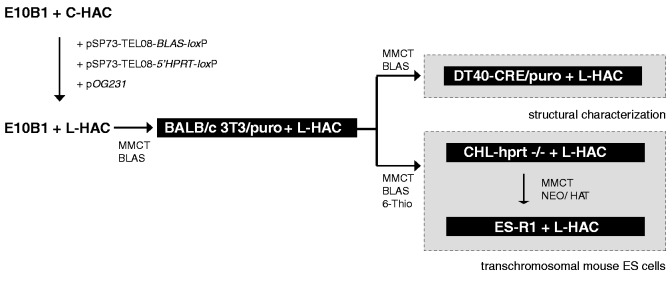
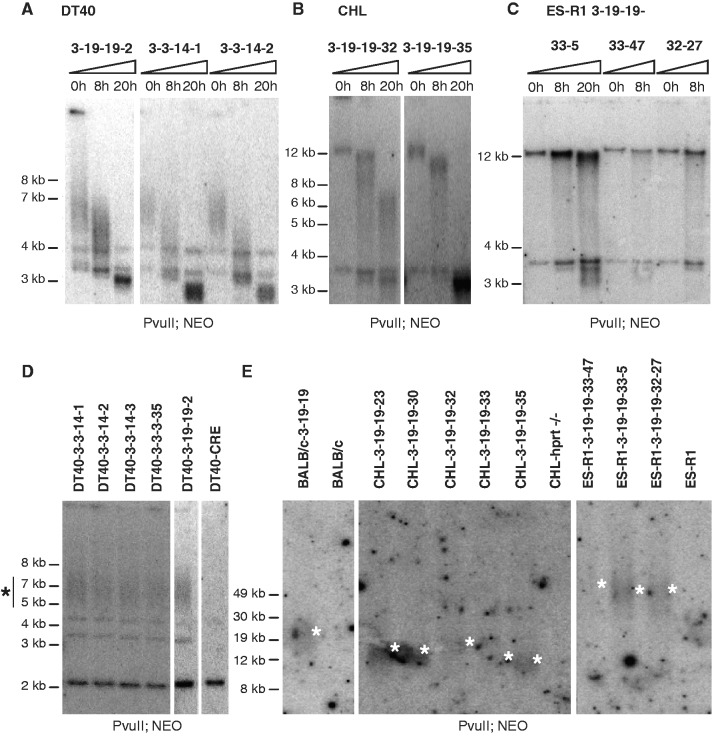
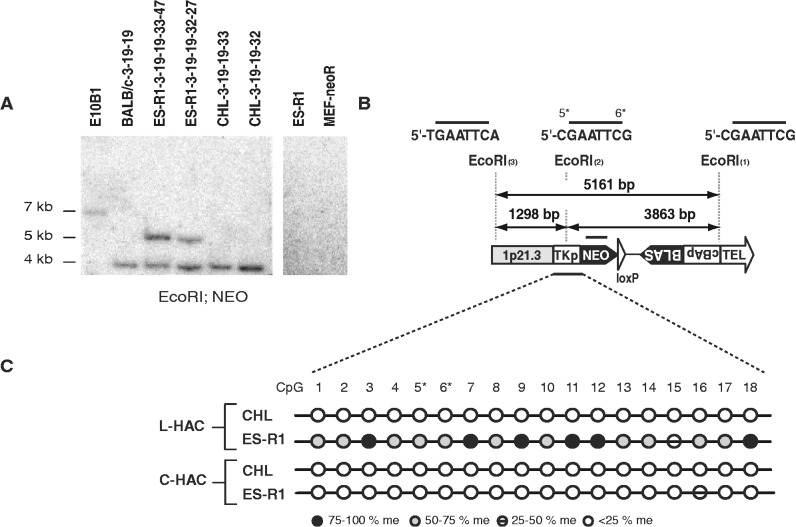
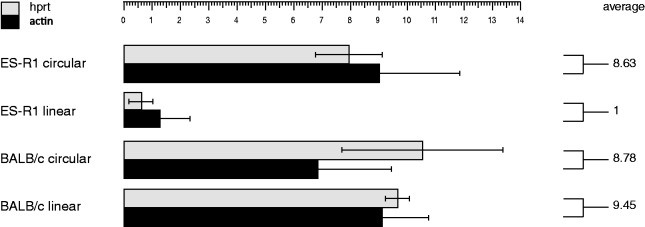
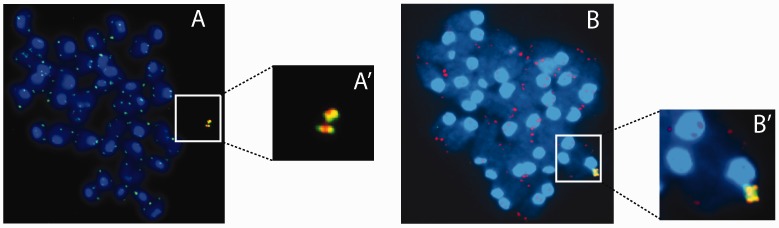
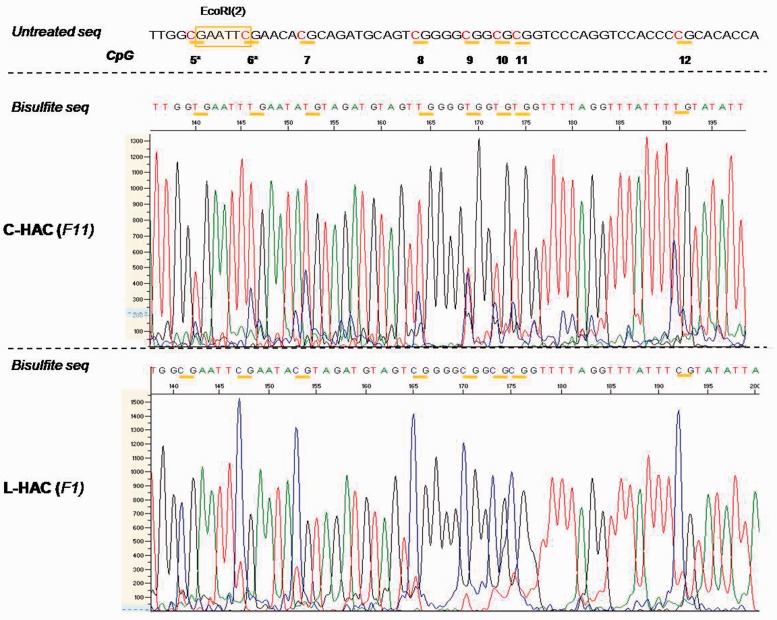
Telomere position effect (TPE) is the influence of telomeres on subtelomeric epigenetic marks and gene expression. Previous studies suggested that TPE depends on genetic background. As these analyses were performed on different chromosomes, cell types and species, it remains unclear whether TPE represents a chromosome-rather than genetic background-specific regulation. We describe the development of a Linear Human Artificial Chromosome (L-HAC) as a new tool for telomere studies. The L-HAC was generated through the Cre-loxP-mediated addition of telomere ends to an existing circular HAC (C-HAC). As it can be transferred to genetically distinct cell lines and animal models the L-HAC enables the study of TPE in an unprecedented manner. The HAC was relocated to four telomerase-positive cell lines via microcell-mediated chromosome transfer and subsequently to mice via blastocyst injection of L-HAC(+)-ES-cells. We could show consistent genetic background-dependent adaptation of telomere length and telomere-associated de novo subtelomeric DNA methylation in mouse ES-R1 cells as well as in mice. Expression of the subtelomeric neomycin gene was inversely correlated with telomere length and subtelomeric methylation. We thus provide a new tool for functional telomere studies and provide strong evidence that telomere length, subtelomeric chromatin marks and expression of subtelomeric genes are genetic background dependent.
Figures
Similar articles
-
A human artificial chromosome recapitulates the metabolism of native telomeres in mammalian cells.PLoS One. 2014 Feb 18;9(2):e88530. doi: 10.1371/journal.pone.0088530. eCollection 2014. PLoS One. 2014. PMID: 24558398 Free PMC article.
-
Systematic Analysis of Compounds Specifically Targeting Telomeres and Telomerase for Clinical Implications in Cancer Therapy.Cancer Res. 2018 Nov 1;78(21):6282-6296. doi: 10.1158/0008-5472.CAN-18-0894. Epub 2018 Aug 30. Cancer Res. 2018. PMID: 30166419 Free PMC article.
-
DNA methylation mediated up-regulation of TERRA non-coding RNA is coincident with elongated telomeres in the human placenta.Mol Hum Reprod. 2016 Nov;22(11):791-799. doi: 10.1093/molehr/gaw053. Epub 2016 Sep 7. Mol Hum Reprod. 2016. PMID: 27604461
-
Human subtelomeric DNA methylation: regulation and roles in telomere function.Curr Opin Genet Dev. 2020 Feb;60:9-16. doi: 10.1016/j.gde.2020.02.004. Epub 2020 Feb 25. Curr Opin Genet Dev. 2020. PMID: 32109830 Review.
-
Developing de novo human artificial chromosomes in embryonic stem cells using HSV-1 amplicon technology.Chromosome Res. 2015 Feb;23(1):105-10. doi: 10.1007/s10577-014-9456-2. Chromosome Res. 2015. PMID: 25657030 Free PMC article. Review.
Cited by
-
A human artificial chromosome recapitulates the metabolism of native telomeres in mammalian cells.PLoS One. 2014 Feb 18;9(2):e88530. doi: 10.1371/journal.pone.0088530. eCollection 2014. PLoS One. 2014. PMID: 24558398 Free PMC article.
-
SORBS2 transcription is activated by telomere position effect-over long distance upon telomere shortening in muscle cells from patients with facioscapulohumeral dystrophy.Genome Res. 2015 Dec;25(12):1781-90. doi: 10.1101/gr.190660.115. Epub 2015 Sep 10. Genome Res. 2015. PMID: 26359233 Free PMC article.
-
Viewing Nuclear Architecture through the Eyes of Nocturnal Mammals.Trends Cell Biol. 2020 Apr;30(4):276-289. doi: 10.1016/j.tcb.2019.12.008. Epub 2020 Jan 22. Trends Cell Biol. 2020. PMID: 31980345 Free PMC article. Review.
-
Regulation of Gene Expression by Telomere Position Effect.Int J Mol Sci. 2021 Nov 26;22(23):12807. doi: 10.3390/ijms222312807. Int J Mol Sci. 2021. PMID: 34884608 Free PMC article. Review.
-
Involvement of Inheritance in Determining Telomere Length beyond Environmental and Lifestyle Factors.Aging Dis. 2023 Nov 5;15(6):2470-2490. doi: 10.14336/AD.2023.1023. Aging Dis. 2023. PMID: 37962459 Free PMC article. Review.
References
-
- Grimes BR, Monaco ZL. Artificial and engineered chromosomes: developments and prospects for gene therapy. Chromosoma. 2005;114:230–241. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
