

The Haemophilus influenzae Sap transporter mediates bacterium-epithelial cell homeostasis
- PMID: 23071138
- PMCID: PMC3536144
- DOI: 10.1128/IAI.00942-12
The Haemophilus influenzae Sap transporter mediates bacterium-epithelial cell homeostasis
Abstract
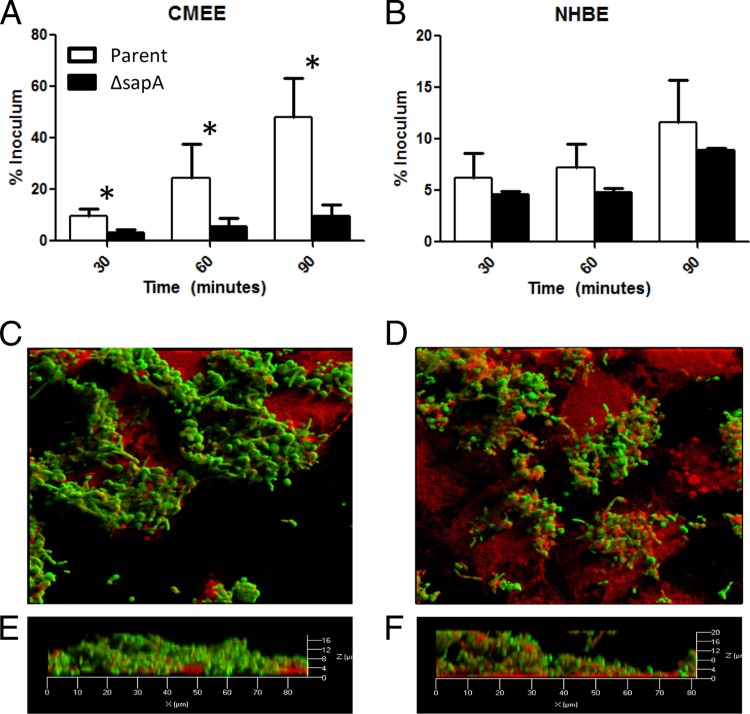
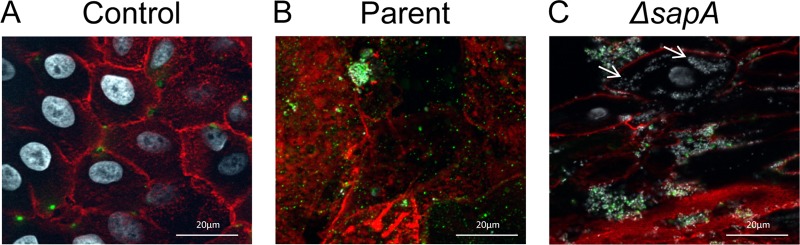
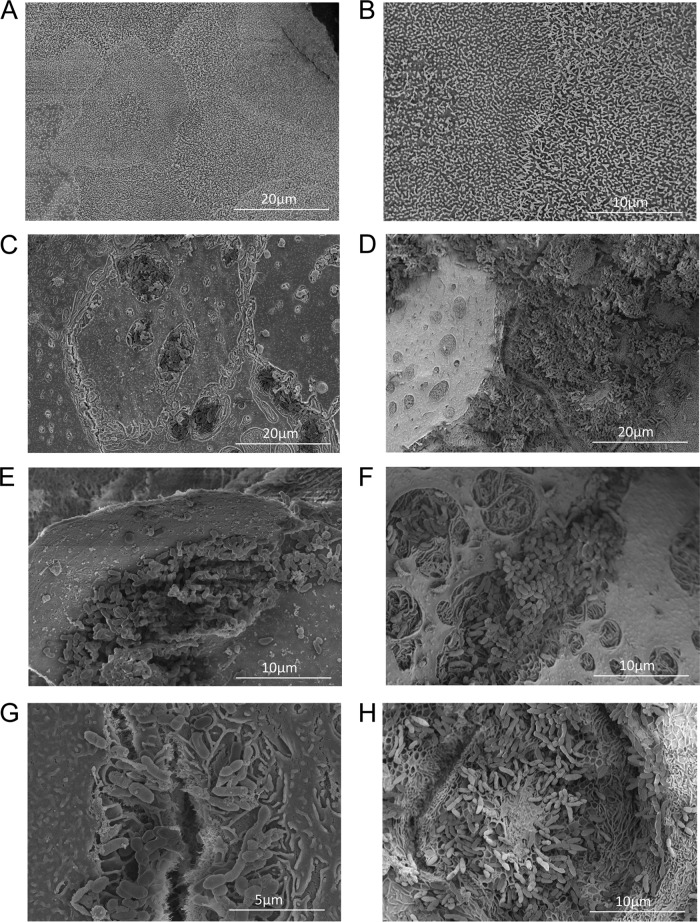
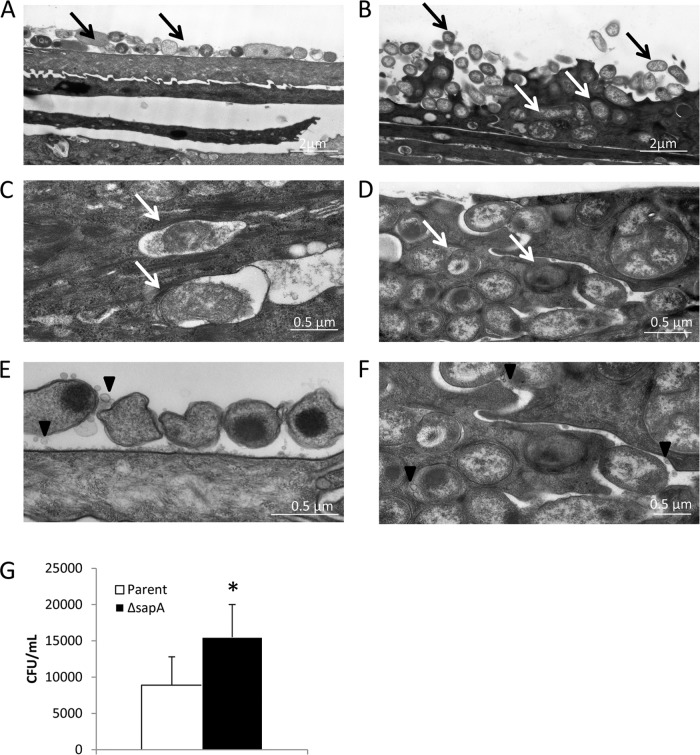
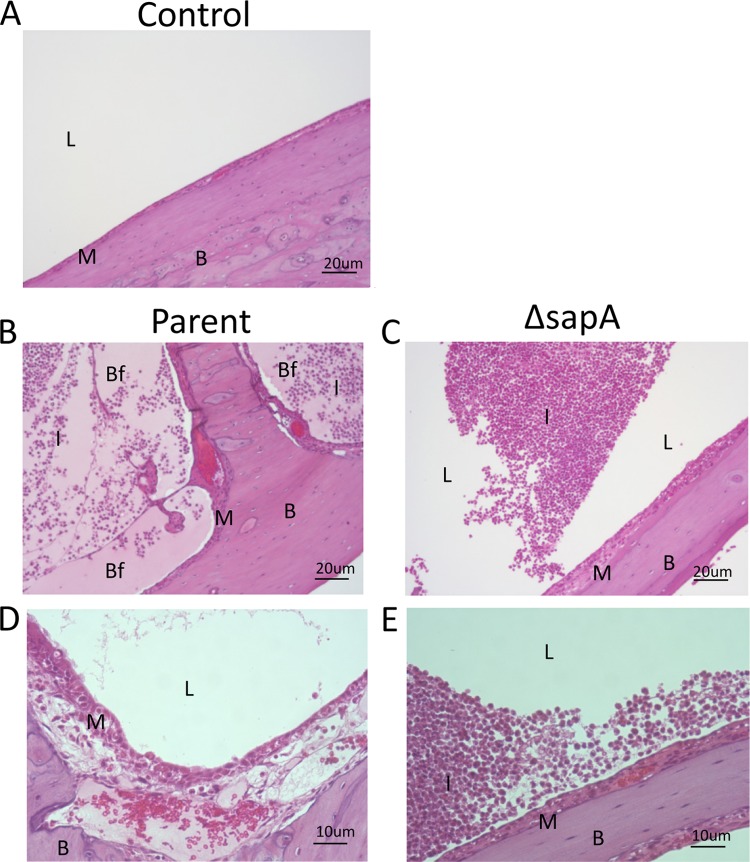
Nontypeable Haemophilus influenzae (NTHI) is a commensal inhabitant of the human nasopharynx and a causative agent of otitis media and other diseases of the upper and lower human airway. During colonization within the host, NTHI must acquire essential nutrients and evade immune attack. We previously demonstrated that the NTHI Sap transporter, an inner membrane protein complex, mediates resistance to antimicrobial peptides and is required for heme homeostasis. We hypothesized that Sap transporter functions are critical for NTHI interaction with the host epithelium and establishment of colonization. Thus, we cocultured the parent or the sapA mutant on polarized epithelial cells grown at an air-liquid interface, as a physiological model of NTHI colonization, to determine the contribution of the Sap transporter to bacterium-host cell interactions. Although SapA-deficient NTHI was less adherent to epithelial cells, we observed a significant increase in invasive bacteria compared to the parent strain. Upon internalization, the sapA mutant appeared free in the cytoplasm, whereas the parent strain was primarily found in endosomes, indicating differential subcellular trafficking. Additionally, we observed reduced inflammatory cytokine production by the epithelium in response to the sapA mutant strain compared to the parental strain. Furthermore, chinchilla middle ears challenged with the sapA mutant demonstrated a decrease in disease severity compared to ears challenged with the parental strain. Collectively, our data suggest that NTHI senses host environmental cues via Sap transporter function to mediate interaction with host epithelial cells. Epithelial cell invasion and modulation of host inflammatory cytokine responses may promote NTHI colonization and access to essential nutrients.
Figures
Similar articles
-
Sap transporter mediated import and subsequent degradation of antimicrobial peptides in Haemophilus.PLoS Pathog. 2011 Nov;7(11):e1002360. doi: 10.1371/journal.ppat.1002360. Epub 2011 Nov 3. PLoS Pathog. 2011. PMID: 22072973 Free PMC article.
-
A mutation in the sap operon attenuates survival of nontypeable Haemophilus influenzae in a chinchilla model of otitis media.Infect Immun. 2005 Jan;73(1):599-608. doi: 10.1128/IAI.73.1.599-608.2005. Infect Immun. 2005. PMID: 15618200 Free PMC article.
-
SapF-mediated heme-iron utilization enhances persistence and coordinates biofilm architecture of Haemophilus.Front Cell Infect Microbiol. 2012 Apr 3;2:42. doi: 10.3389/fcimb.2012.00042. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 22919633 Free PMC article.
-
Developing a nontypeable Haemophilus influenzae (NTHi) vaccine.Vaccine. 2000 Dec 8;19 Suppl 1:S108-15. doi: 10.1016/s0264-410x(00)00288-7. Vaccine. 2000. PMID: 11163473 Review.
-
Impact of protein D-containing pneumococcal conjugate vaccines on non-typeable Haemophilus influenzae acute otitis media and carriage.Expert Rev Vaccines. 2017 Jul;16(7):1-14. doi: 10.1080/14760584.2017.1333905. Epub 2017 Jun 7. Expert Rev Vaccines. 2017. PMID: 28571504 Review.
Cited by
-
Tolerance to Haemophilus influenzae infection in human epithelial cells: Insights from a primary cell-based model.PLoS Pathog. 2024 Jul 11;20(7):e1012282. doi: 10.1371/journal.ppat.1012282. eCollection 2024 Jul. PLoS Pathog. 2024. PMID: 38990812 Free PMC article.
-
Pathogenic Mannheimia haemolytica Invades Differentiated Bovine Airway Epithelial Cells.Infect Immun. 2019 May 21;87(6):e00078-19. doi: 10.1128/IAI.00078-19. Print 2019 Jun. Infect Immun. 2019. PMID: 30962401 Free PMC article.
-
Role of sapA and yfgA in Susceptibility to Antibody-Mediated Complement-Dependent Killing and Virulence of Salmonella enterica Serovar Typhimurium.Infect Immun. 2017 Aug 18;85(9):e00419-17. doi: 10.1128/IAI.00419-17. Print 2017 Sep. Infect Immun. 2017. PMID: 28674031 Free PMC article.
-
In vitro modelling of bacterial pneumonia: a comparative analysis of widely applied complex cell culture models.FEMS Microbiol Rev. 2024 Mar 1;48(2):fuae007. doi: 10.1093/femsre/fuae007. FEMS Microbiol Rev. 2024. PMID: 38409952 Free PMC article. Review.
-
Reprioritization of biofilm metabolism is associated with nutrient adaptation and long-term survival of Haemophilus influenzae.NPJ Biofilms Microbiomes. 2019 Nov 5;5(1):33. doi: 10.1038/s41522-019-0105-6. eCollection 2019. NPJ Biofilms Microbiomes. 2019. PMID: 31700653 Free PMC article.
References
-
- Groeneveld K, van Alphen L, Eijk PP, Visschers G, Jansen HM, Zanen HC. 1990. Endogenous and exogenous reinfections by Haemophilus influenzae in patients with chronic obstructive pulmonary disease: the effect of antibiotic treatment on persistence. J. Infect. Dis. 161:512–517 - PubMed
-
- Klein JO. 1997. Role of nontypeable Haemophilus influenzae in pediatric respiratory tract infections. Pediatr. Infect. Dis. J. 16:S5–8 - PubMed
-
- Murphy TF. 2003. Respiratory infections caused by non-typeable Haemophilus influenzae. Curr. Opin. Infect. Dis. 16:129–134 - PubMed
-
- Murphy TF, Brauer AL, Schiffmacher AT, Sethi S. 2004. Persistent colonization by Haemophilus influenzae in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 170:266–272 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
