

TRPA1 has a key role in the somatic pro-nociceptive actions of hydrogen sulfide
- PMID: 23071662
- PMCID: PMC3469557
- DOI: 10.1371/journal.pone.0046917
TRPA1 has a key role in the somatic pro-nociceptive actions of hydrogen sulfide
Abstract
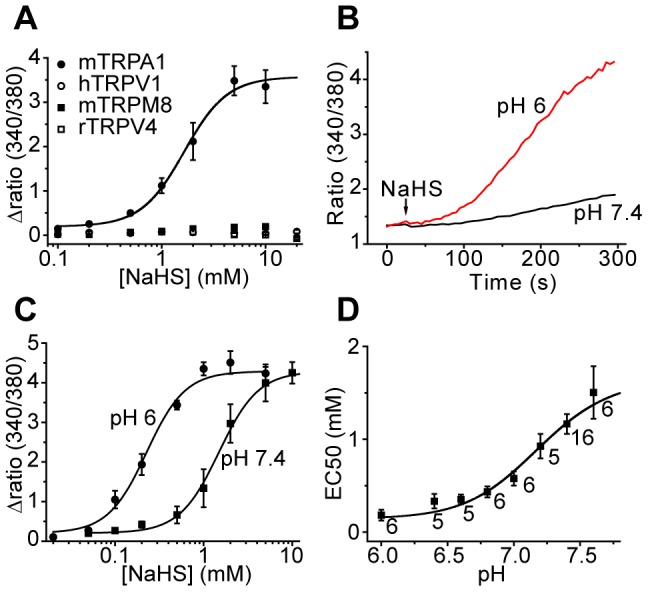
Hydrogen sulfide (H(2)S), which is produced endogenously from L-cysteine, is an irritant with pro-nociceptive actions. We have used measurements of intracellular calcium concentration, electrophysiology and behavioral measurements to show that the somatic pronociceptive actions of H(2)S require TRPA1. A H(2)S donor, NaHS, activated TRPA1 expressed in CHO cells and stimulated DRG neurons isolated from Trpa1(+/+) but not Trpa1(-/-) mice. TRPA1 activation by NaHS was pH dependent with increased activity at acidic pH. The midpoint of the relationship between NaHS EC(50) values and external pH was pH 7.21, close to the expected dissociation constant for H(2)S (pK(a) 7.04). NaHS evoked single channel currents in inside-out and cell-attached membrane patches consistent with an intracellular site of action. In behavioral experiments, intraplantar administration of NaHS and L-cysteine evoked mechanical and cold hypersensitivities in Trpa1(+/+) but not in Trpa1(-/-) mice. The sensitizing effects of L-cysteine in wild-type mice were inhibited by a cystathionine β-synthase inhibitor, D,L-propargylglycine (PAG), which inhibits H(2)S formation. Mechanical hypersensitivity evoked by intraplantar injections of LPS was prevented by PAG and the TRPA1 antagonist AP-18 and was absent in Trpa1(-/-) mice, indicating that H(2)S mediated stimulation of TRPA1 is necessary for the local pronociceptive effects of LPS. The pro-nociceptive effects of intraplantar NaHS were retained in Trpv1(-/-) mice ruling out TRPV1 as a molecular target. In behavioral studies, NaHS mediated sensitization was also inhibited by a T-type calcium channel inhibitor, mibefradil. In contrast to the effects of NaHS on somatic sensitivity, intracolonic NaHS administration evoked similar nociceptive effects in Trpa1(+/+) and Trpa1(-/-) mice, suggesting that the visceral pro-nociceptive effects of H(2)S are independent of TRPA1. In electrophysiological studies, the depolarizing actions of H(2)S on isolated DRG neurons were inhibited by AP-18, but not by mibefradil indicating that the primary excitatory effect of H(2)S on DRG neurons is TRPA1 mediated depolarization.
Conflict of interest statement
Figures








References
-
- Szabo C (2007) Hydrogen sulphide and its therapeutic potential. Nat Rev Drug Discov 6: 917–935. - PubMed
-
- Kimura H (2010) Hydrogen Sulfide: From Brain to Gut. Antioxid Redox Signal - PubMed
-
- Distrutti E, Sediari L, Mencarelli A, Renga B, Orlandi S, et al. (2006) Evidence that hydrogen sulfide exerts antinociceptive effects in the gastrointestinal tract by activating KATP channels. J Pharmacol Exp Ther 316: 325–335. - PubMed
-
- Distrutti E, Sediari L, Mencarelli A, Renga B, Orlandi S, et al. (2006) 5-Amino-2-hydroxybenzoic acid 4-(5-thioxo-5H-[1,2]dithiol-3yl)-phenyl ester (ATB-429), a hydrogen sulfide-releasing derivative of mesalamine, exerts antinociceptive effects in a model of postinflammatory hypersensitivity. J Pharmacol Exp Ther 319: 447–458. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
