

Knockout of multiple Arabidopsis cation/H(+) exchangers suggests isoform-specific roles in metal stress response, germination and seed mineral nutrition
- PMID: 23071810
- PMCID: PMC3470555
- DOI: 10.1371/journal.pone.0047455
Knockout of multiple Arabidopsis cation/H(+) exchangers suggests isoform-specific roles in metal stress response, germination and seed mineral nutrition
Abstract
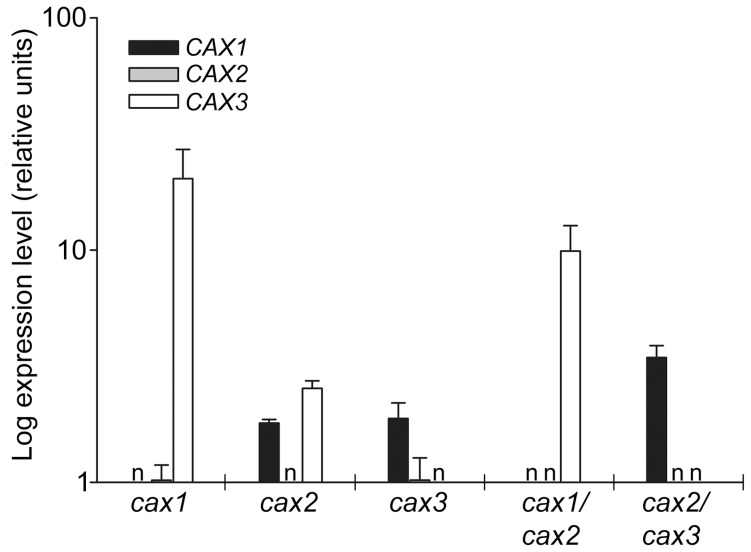
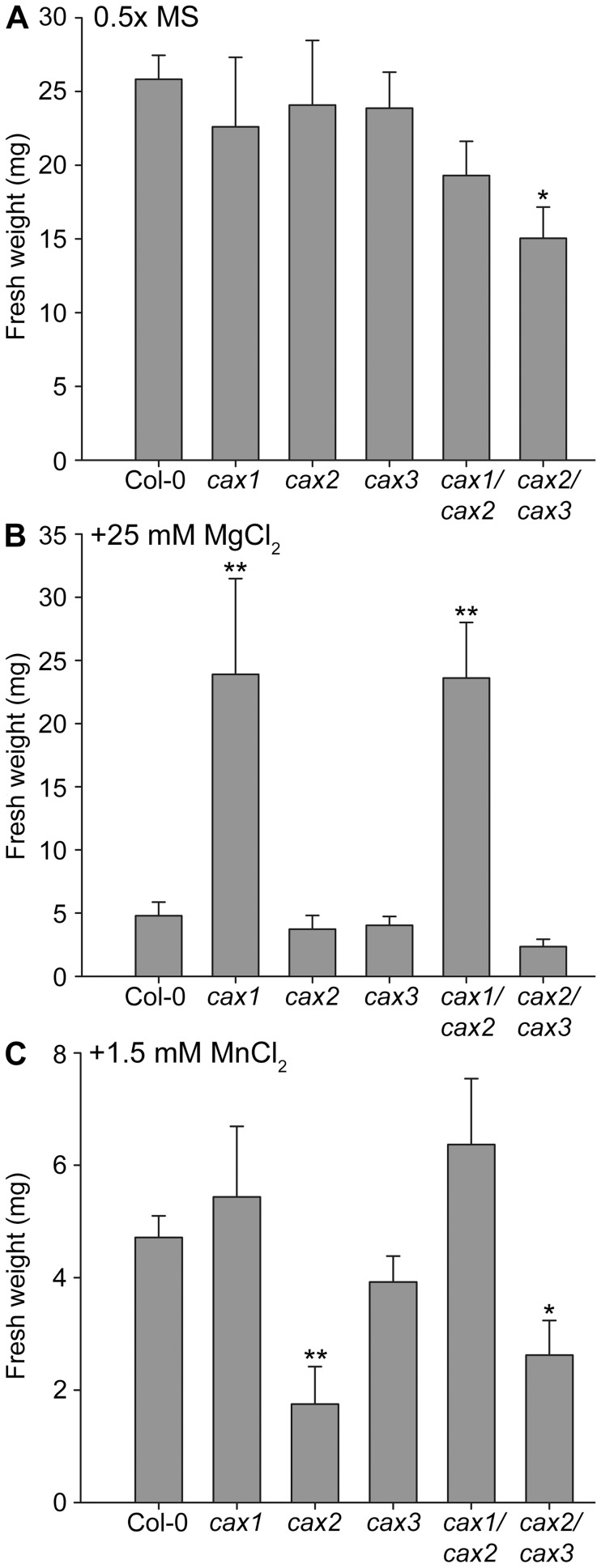
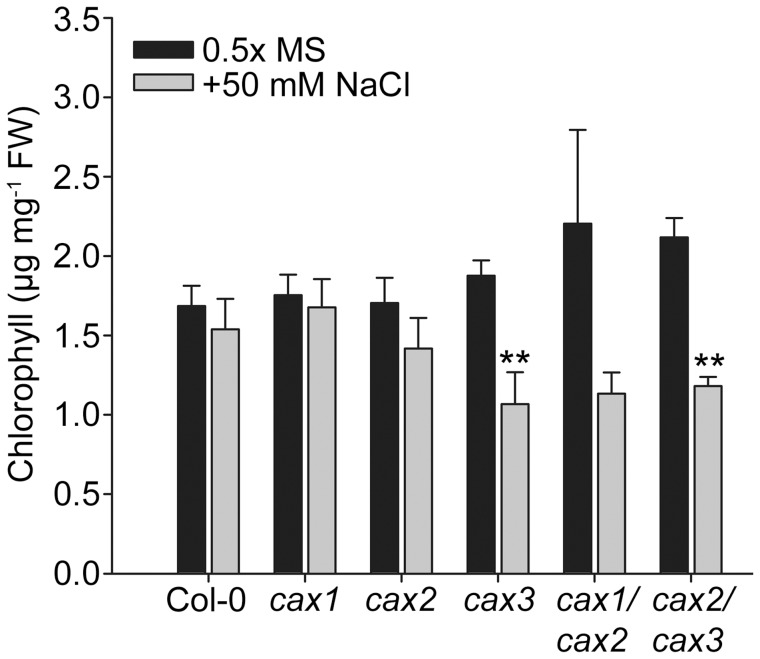
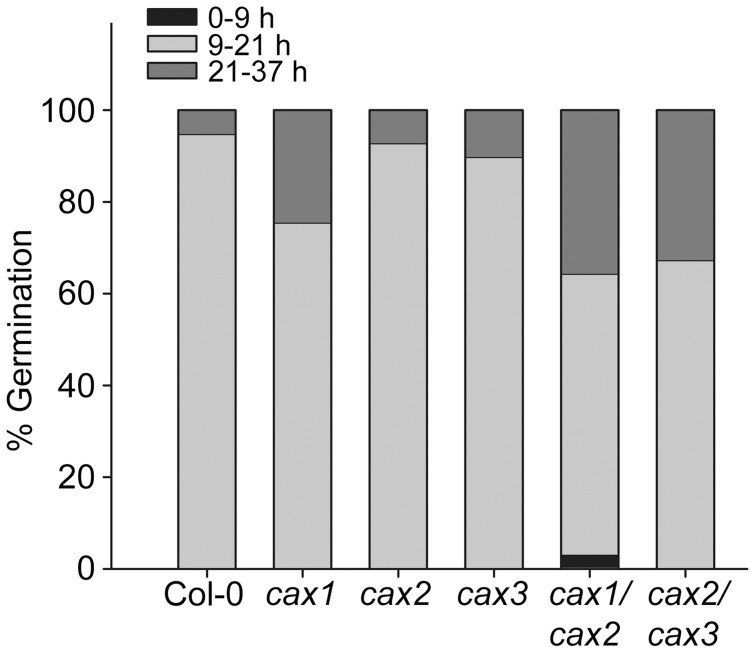
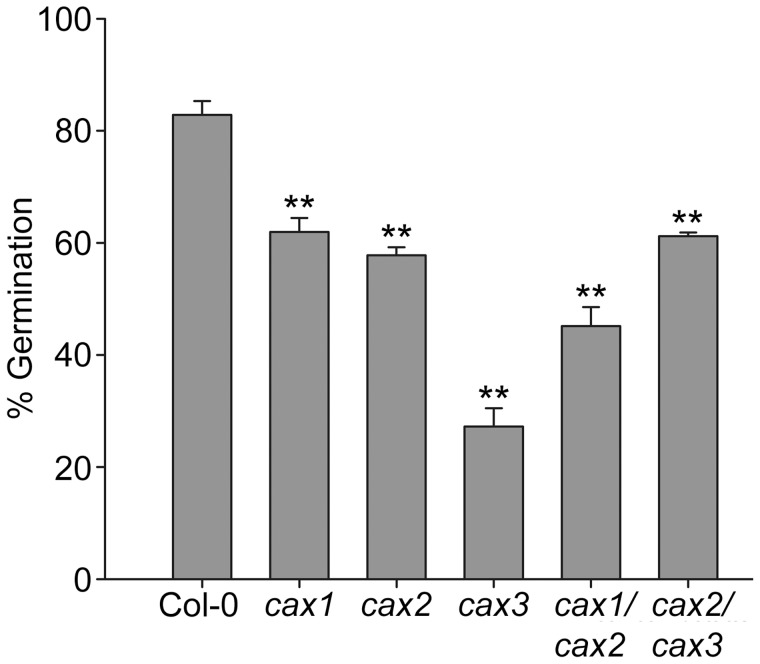
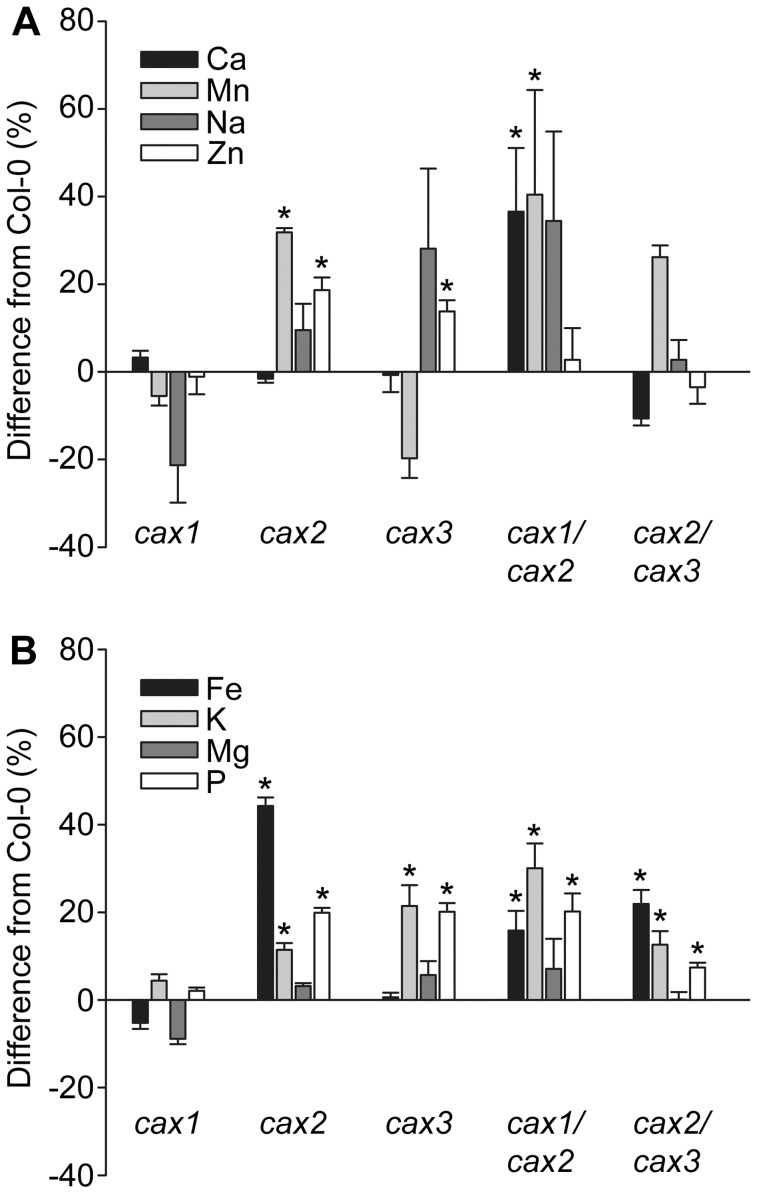
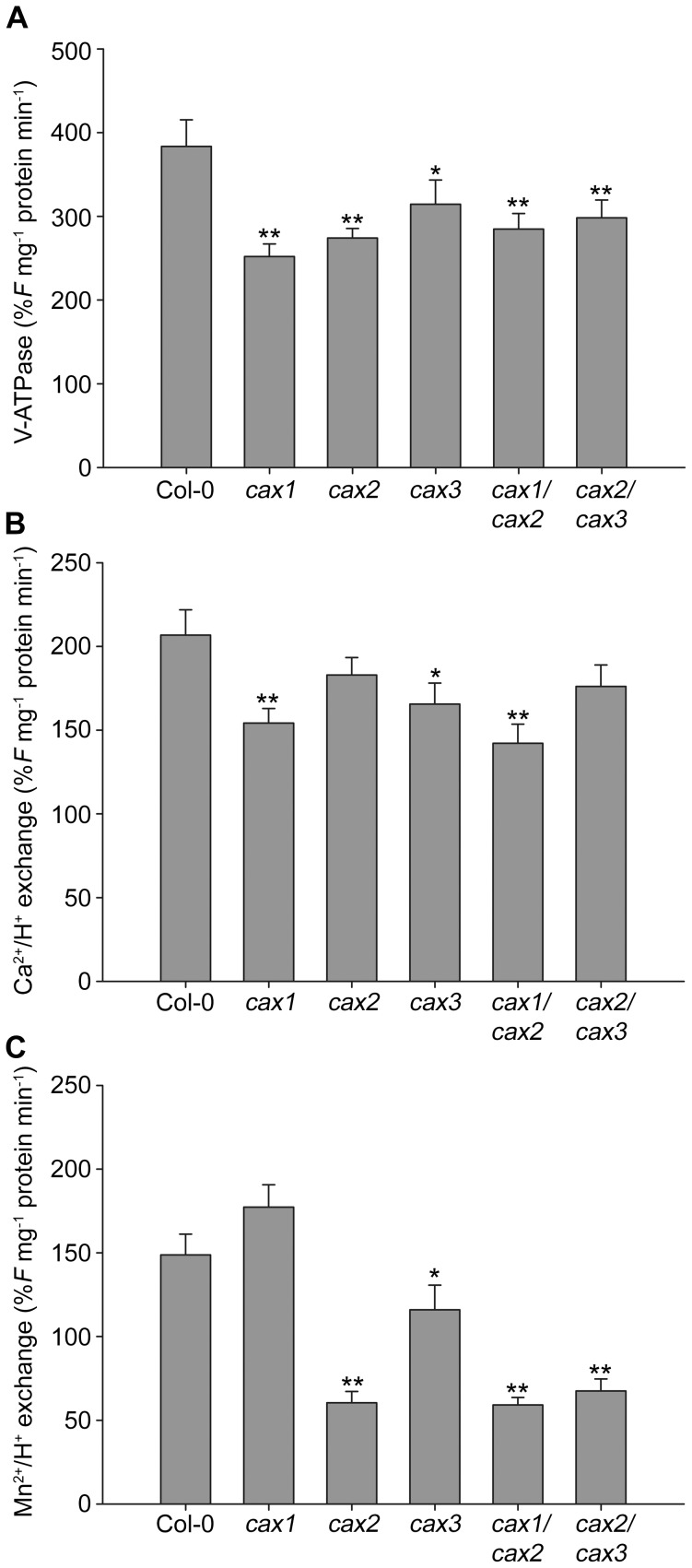
Cation/H(+) exchangers encoded by CAX genes play an important role in the vacuolar accumulation of metals including Ca(2+) and Mn(2+). Arabidopsis thaliana CAX1 and CAX3 have been previously shown to differ phylogenetically from CAX2 but the physiological roles of these different transporters are still unclear. To examine the functions and the potential of redundancy between these three cation transporters, cax1/cax2 and cax2/cax3 double knockout mutants were generated and compared with wild type and cax single knockouts. These double mutants had equivalent metal stress responses to single cax mutants. Both cax1 and cax1/cax2 had increased tolerance to Mg stress, while cax2 and cax2/cax3 both had increased sensitivity to Mn stress. The cax1/cax2 and cax2/cax3 mutants did not exhibit the deleterious developmental phenotypes previously seen with the cax1/cax3 mutant. However, these new double mutants did show alterations in seed germination, specifically a delay in germination time. These alterations correlated with changes in nutrient content within the seeds of the mutants, particularly the cax1/cax2 mutant which had significantly higher seed content of Ca and Mn. This study indicates that the presence of these Arabidopsis CAX transporters is important for normal germination and infers a role for CAX proteins in metal homeostasis within the seed.
Conflict of interest statement
Figures
References
-
- Colangelo EP, Guerinot ML (2006) Put the metal to the petal: metal uptake and transport throughout plants. Curr Opin Plant Biol 9: 322–330. - PubMed
-
- Puig S, Penarrubia L (2009) Placing metal micronutrients in context: transport and distribution in plants. Curr Opin Plant Biol 12: 299–306. - PubMed
-
- Karley AJ, White PJ (2009) Moving cationic minerals to edible tissues: potassium, magnesium, calcium. Curr Opin Plant Biol 12: 291–298. - PubMed
-
- Manohar M, Shigaki T, Hirschi KD (2011) Plant cation/H+ exchangers (CAXs): biological functions and genetic manipulations. Plant Biol 13: 561–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
