

Herpes simplex virus γ34.5 interferes with autophagosome maturation and antigen presentation in dendritic cells
- PMID: 23073763
- PMCID: PMC3470650
- DOI: 10.1128/mBio.00267-12
Herpes simplex virus γ34.5 interferes with autophagosome maturation and antigen presentation in dendritic cells
Abstract
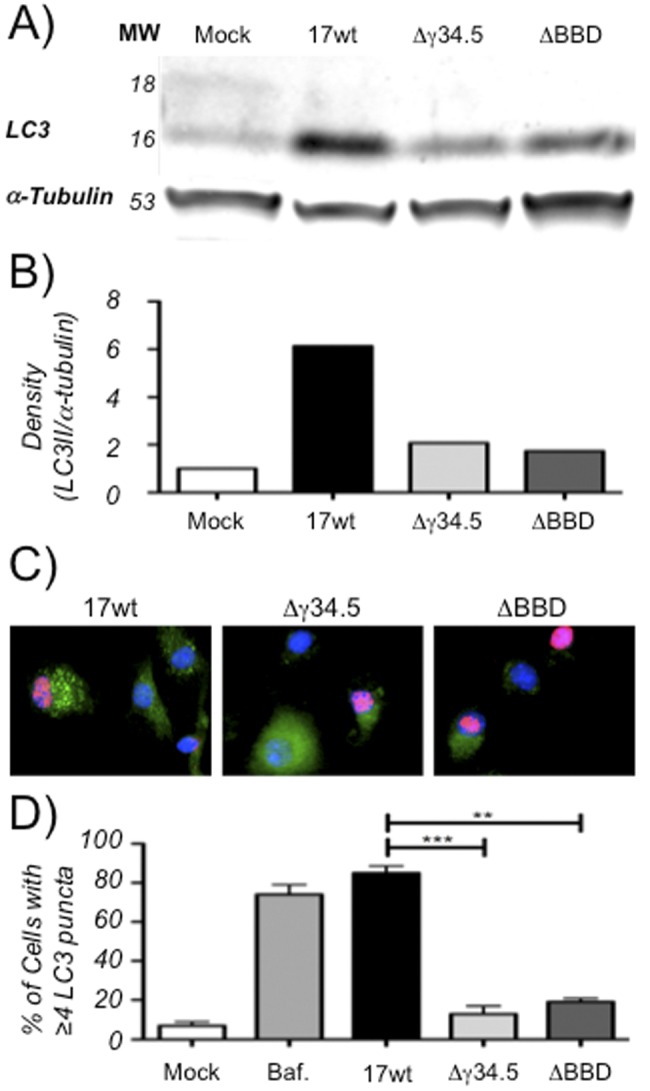
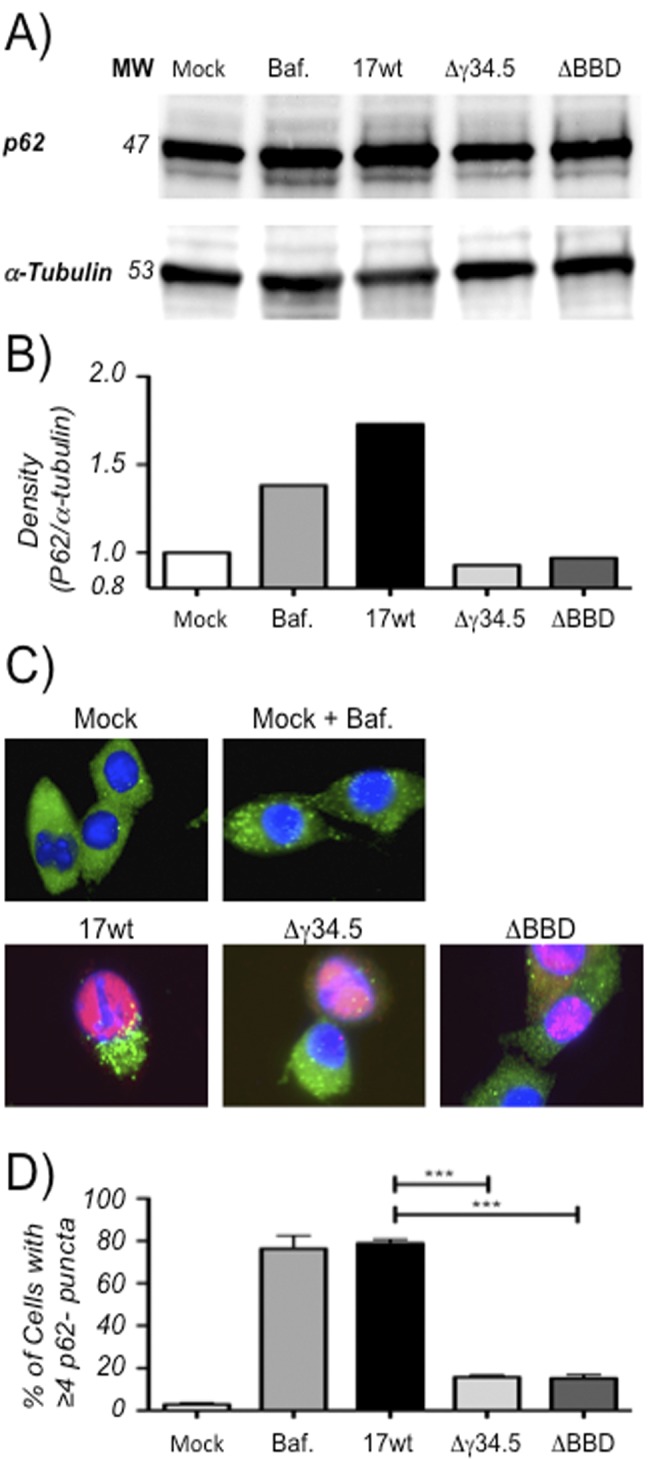
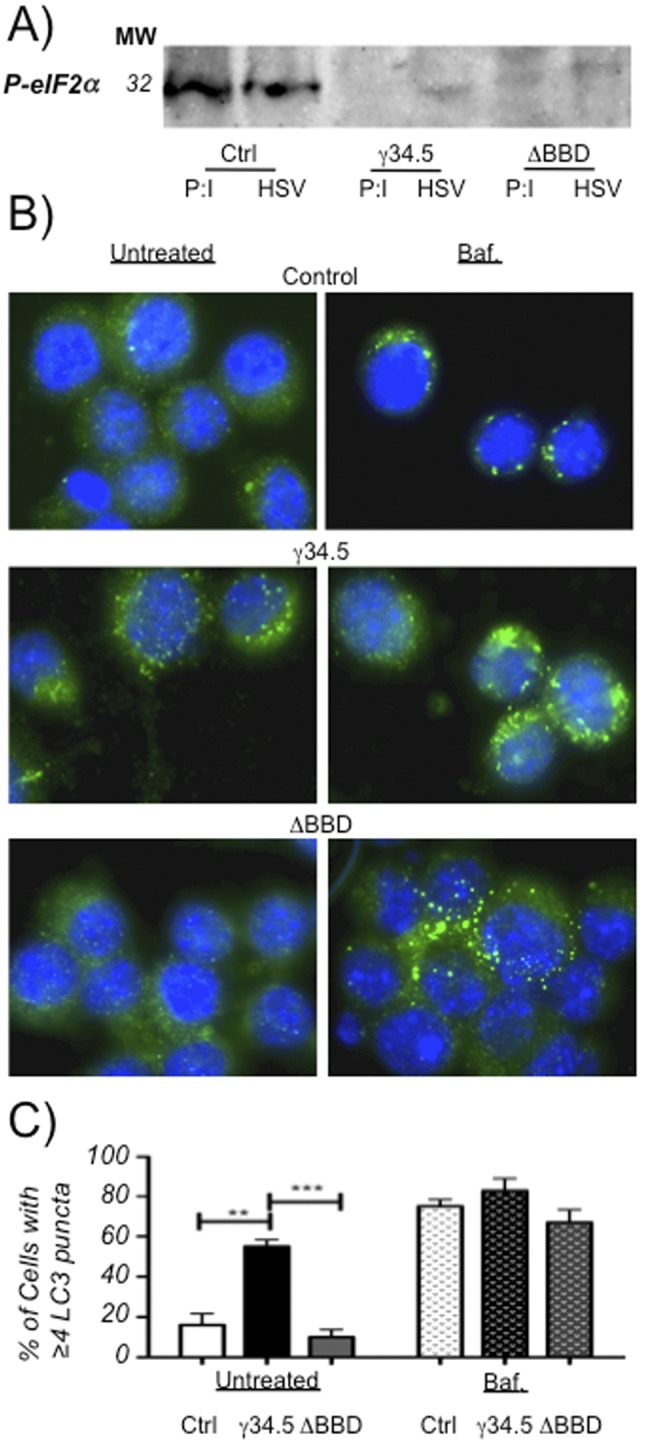
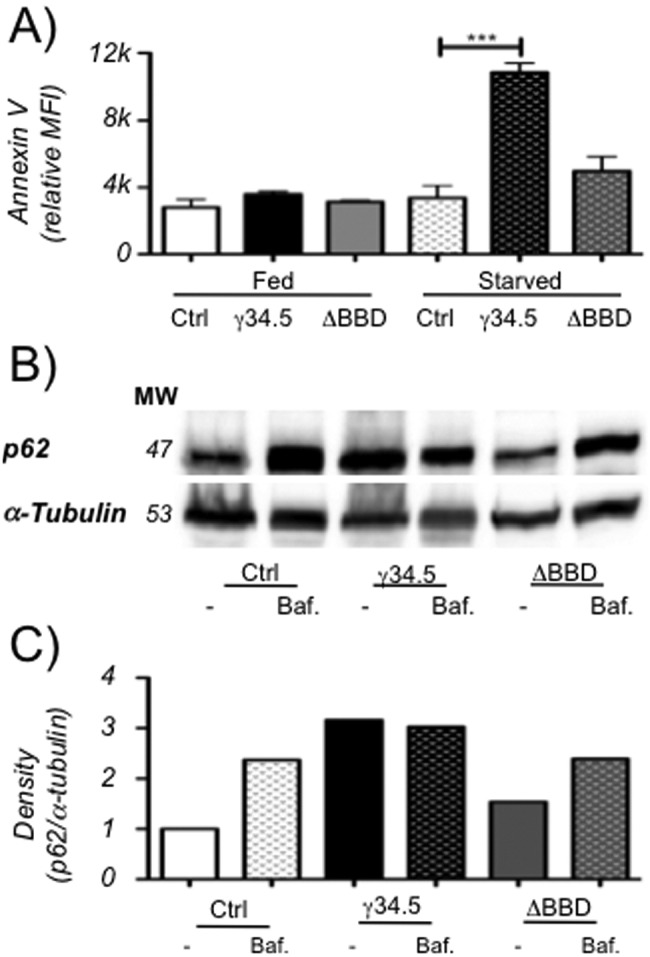
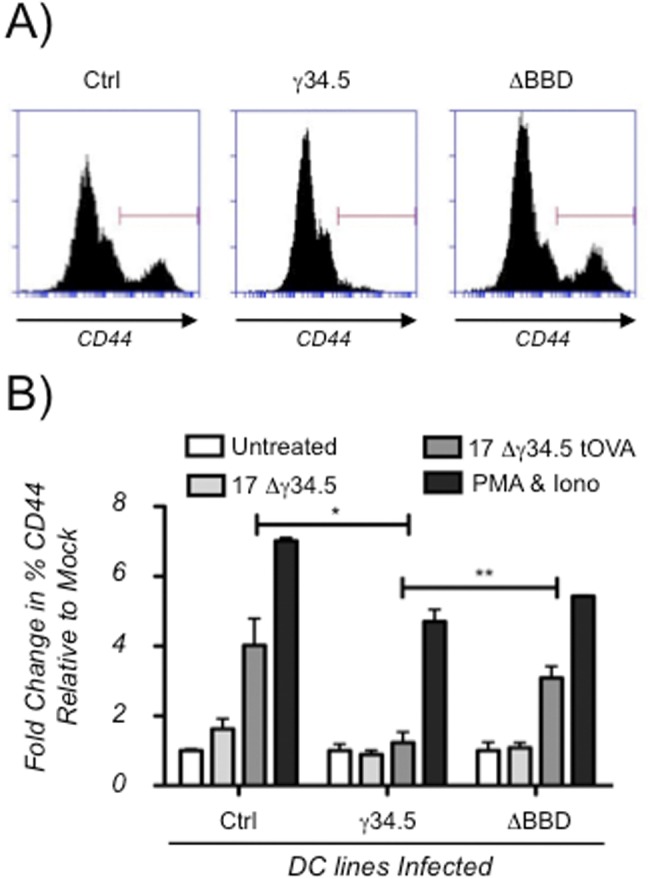
The cellular autophagy response induced by herpes simplex virus 1 (HSV-1) is countered by the viral γ34.5 protein. γ34.5 modulates autophagy by binding to the host autophagy protein Beclin-1 and through this binding inhibits the formation of autophagosomes in fibroblasts and neurons. In contrast, in this study dendritic cells (DCs) infected with HSV-1 showed an accumulation of autophagosomes and of the long-lived protein p62. No such accumulations were observed in DCs infected with a γ34.5-null virus or a virus lacking the Beclin-binding domain (BBD) of γ34.5. To explore this further, we established stably transduced DC lines to show that γ34.5 expression alone induced autophagosome accumulation yet prevented p62 degradation. In contrast, DCs expressing a BBD-deleted mutant of γ34.5 were unable to modulate autophagy. DCs expressing γ34.5 were less capable of stimulating T-cell activation and proliferation in response to intracellular antigens, demonstrating an immunological consequence of inhibiting autophagy. Taken together, these data show that in DCs, γ34.5 antagonizes the maturation of autophagosomes and T cell activation in a BBD-dependent manner, illustrating a unique interface between HSV and autophagy in antigen-presenting cells. IMPORTANCE Herpes simplex virus 1 (HSV-1) is a highly prevalent pathogen causing widespread morbidity and some mortality. HSV infections are lifelong, and there are no vaccines or antivirals to cure HSV infections. The ability of HSV to modulate host immunity is critical for its virulence. HSV inhibits host autophagy, a pathway with importance in many areas of health and disease. Autophagy is triggered by many microbes, some of which harness autophagy for replication; others evade autophagy or prevent it from occurring. Autophagy is critical for host defense, either by directly degrading the invading pathogen ("xenophagy") or by facilitating antigen presentation to T cells. In this study, we show that HSV manipulates autophagy through an unsuspected mechanism with a functional consequence of reducing T cell stimulation. These data further our understanding of how HSV evades host immunity to persist for the lifetime of its host, facilitating its spread in the human population.
Figures
References
-
- Samuel CE, Brody MS. 1990. Biosynthesis of reovirus-specified polypeptides. 2-Aminopurine increases the efficiency of translation of reovirus s1 mRNA but not s4 mRNA in transfected cells. Virology 176:106–113 - PubMed
-
- Chou J, Chen JJ, Gross M, Roizman B. 1995. Association of a M(r) 90,000 phosphoprotein with protein kinase PKR in cells exhibiting enhanced phosphorylation of translation initiation factor eIF-2 alpha and premature shutoff of protein synthesis after infection with gamma 134.5- mutants of herpes simplex virus 1. Proc. Natl. Acad. Sci. U. S. A. 92:10516–10520 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
