

Osteopontin increases heme oxygenase-1 expression and subsequently induces cell migration and invasion in glioma cells
- PMID: 23074199
- PMCID: PMC3480271
- DOI: 10.1093/neuonc/nos262
Osteopontin increases heme oxygenase-1 expression and subsequently induces cell migration and invasion in glioma cells
Abstract
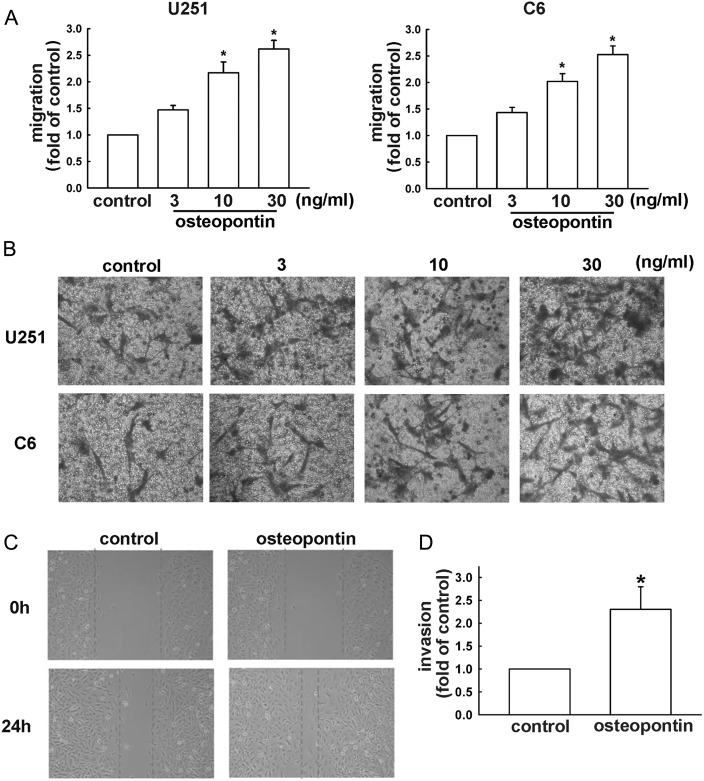
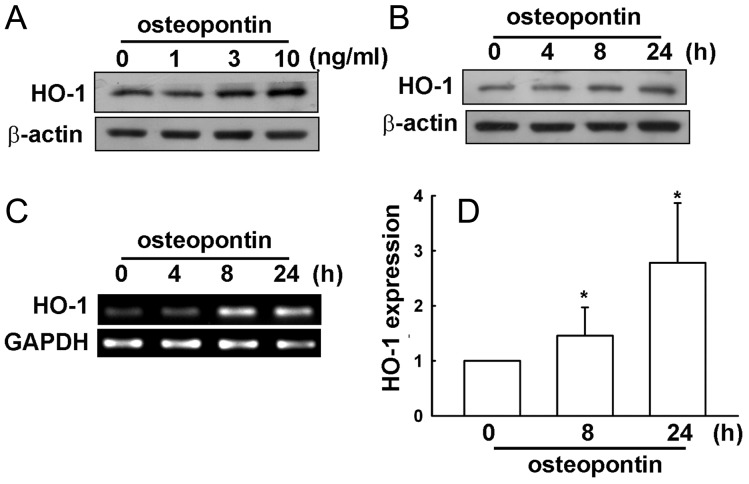
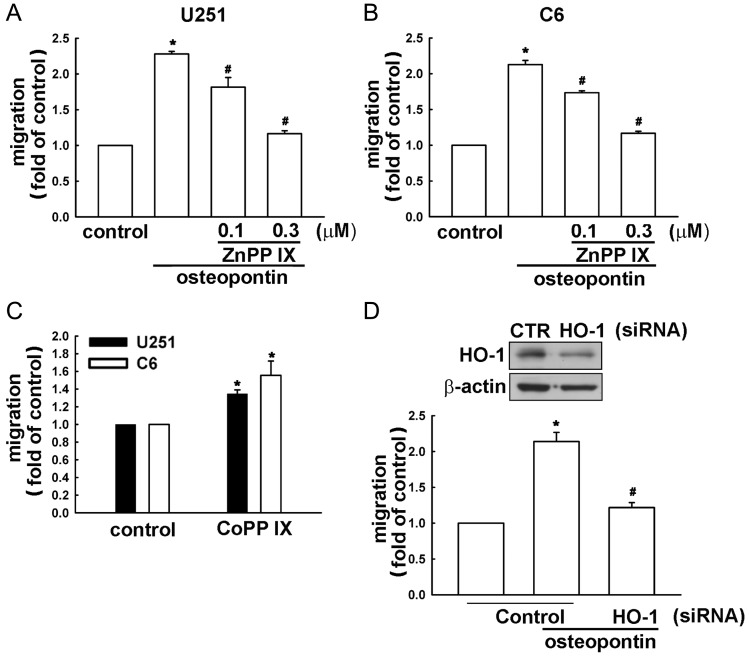
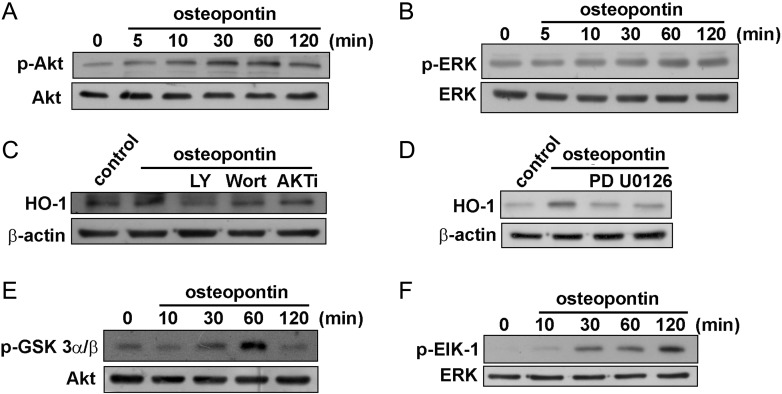
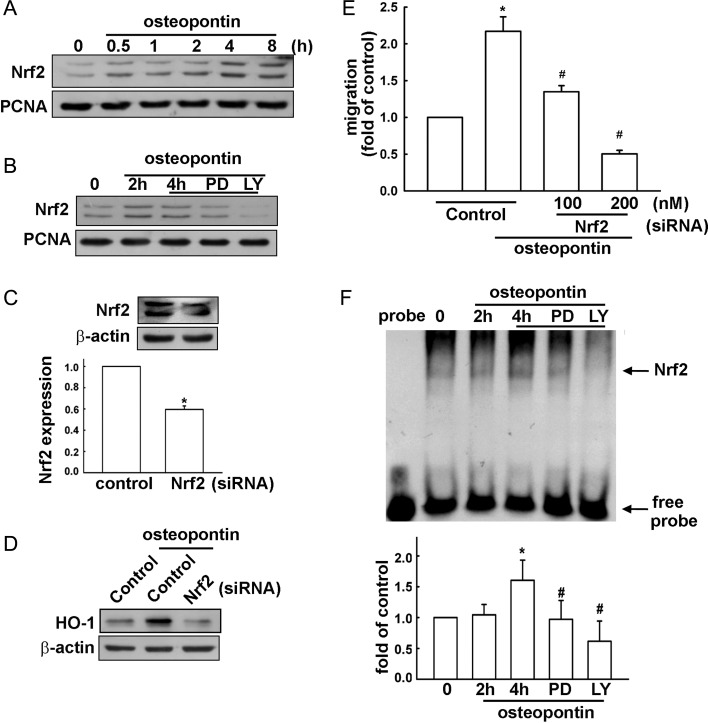
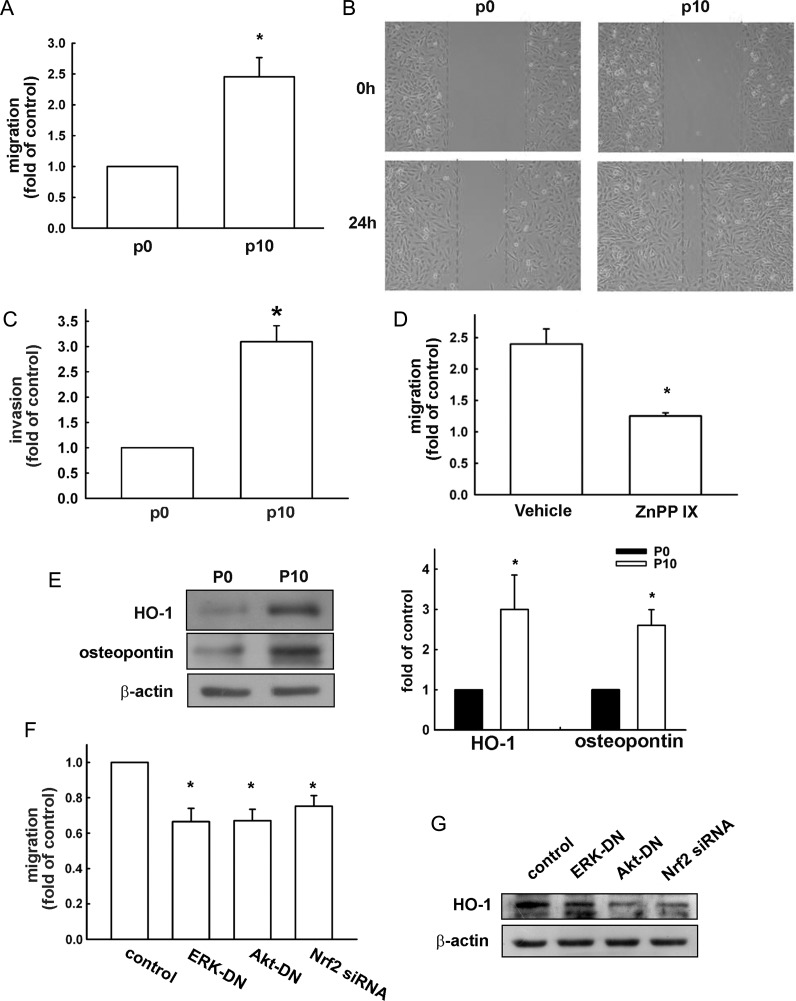
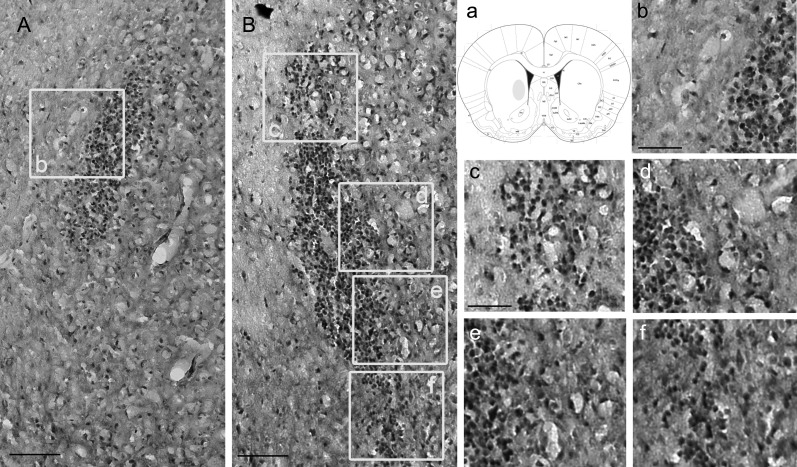
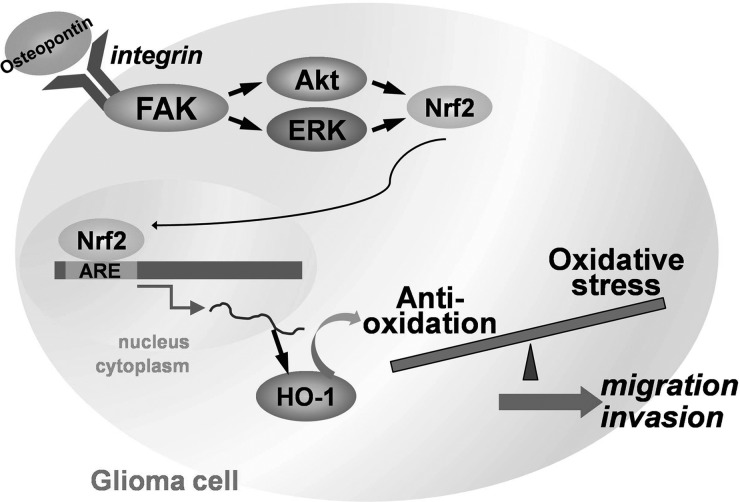
Malignant gliomas are associated with high morbidity and mortality because they are highly invasive into surrounding brain tissue, making complete surgical resection impossible. Osteopontin is abundantly expressed in the brain and is involved in cell adhesion, migration, and invasion. The aim of the present study was to investigate the mechanisms of glioma cell migration. Migration and invasion activity were determined by transwell and wound-healing assays. Gene and protein expressions were analyzed by reverse transcription-PCR, real time-PCR, and Western blotting. Nrf2-DNA binding activity was determined by electrophoretic mobility shift assay. Establishment of migration-prone sublines were performed to select highly migratory glioma. An intracranial xenograft mouse model was used for the in vivo study. Application of recombinant human osteopontin enhanced the migration of glioma cells. Expression of heme oxygenase (HO)-1 mRNA and protein also increased in response to osteopontin stimulation. Osteopontin-induced increase in cell migration was antagonized by HO-1 inhibitor or HO-1 small interfering (si)RNA. Osteopontin-mediated HO-1 expression was reduced by treatment with MEK/ERK and phosphatidylinositol 3-kinase/Akt inhibitors, as well as siRNA against Nrf2. Furthermore, osteopontin stimulated Nrf2 accumulation in the nucleus and increased Nrf2-DNA binding activity. In migration-prone sublines, cells with greater migration ability had higher osteopontin and HO-1 expression, and zinc protoporphyrin IX treatment could effectively reduce the enhanced migration ability. In an intracranial xenograft mouse model, transplantation of migration-prone subline cells exhibited higher cell migration than parental tumor cells. These results indicate that osteopontin activates Nrf2 signaling, resulting in enhanced HO-1 expression and cell migration in glioma cells.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
