

Phosphoenolpyruvate carboxykinase and glucose-6-phosphatase are required for steroidogenesis in testicular Leydig cells
- PMID: 23074219
- PMCID: PMC3516735
- DOI: 10.1074/jbc.M112.421552
Phosphoenolpyruvate carboxykinase and glucose-6-phosphatase are required for steroidogenesis in testicular Leydig cells
Abstract
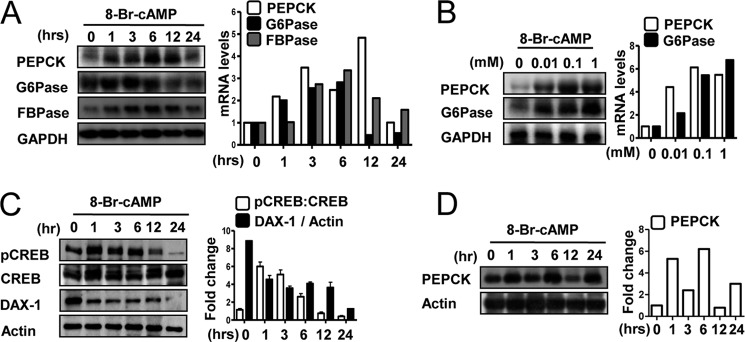
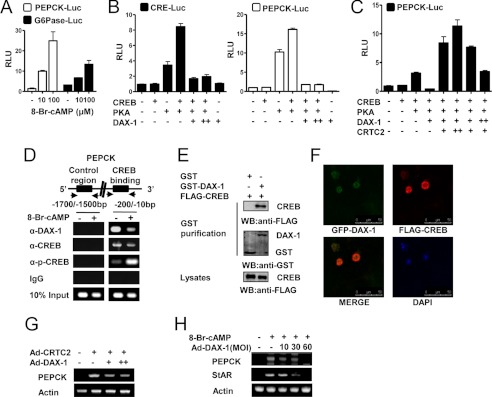
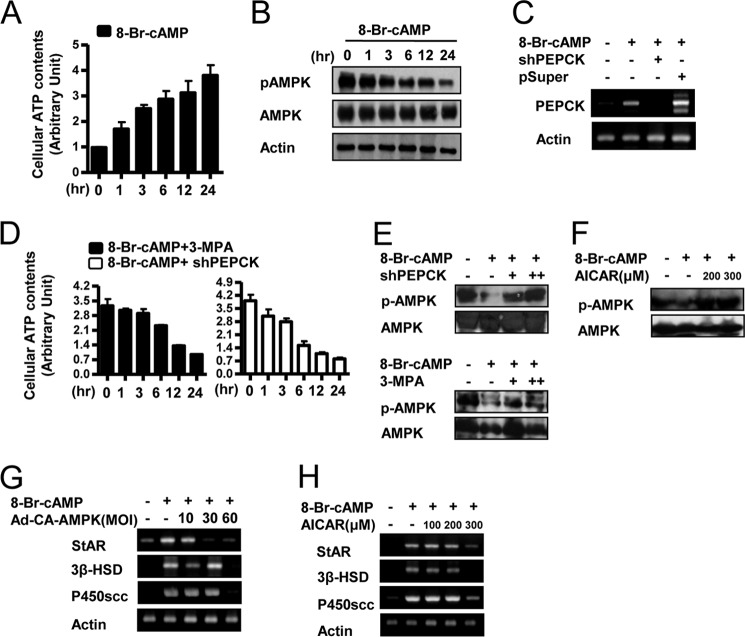
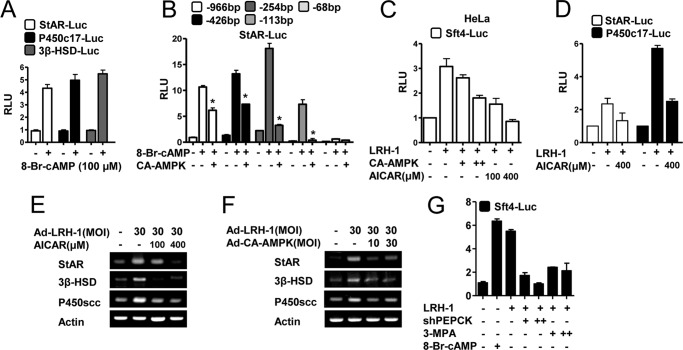
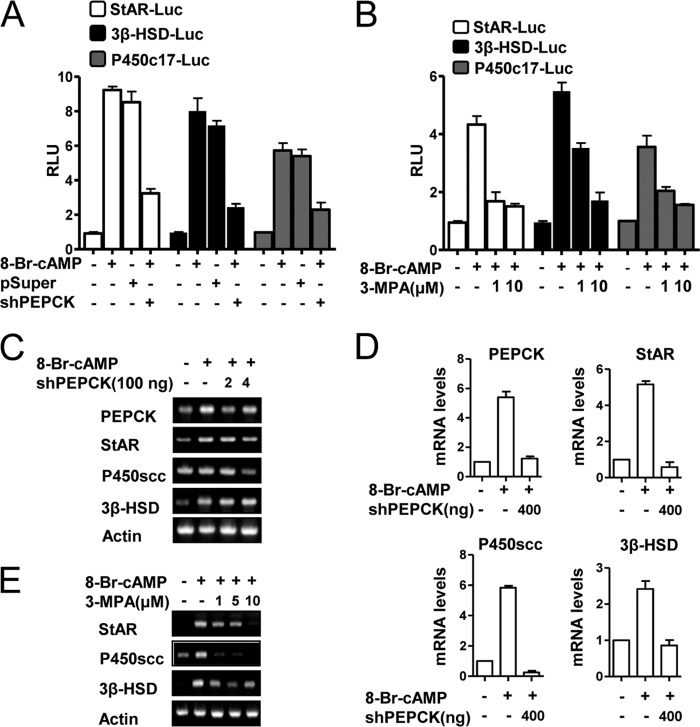
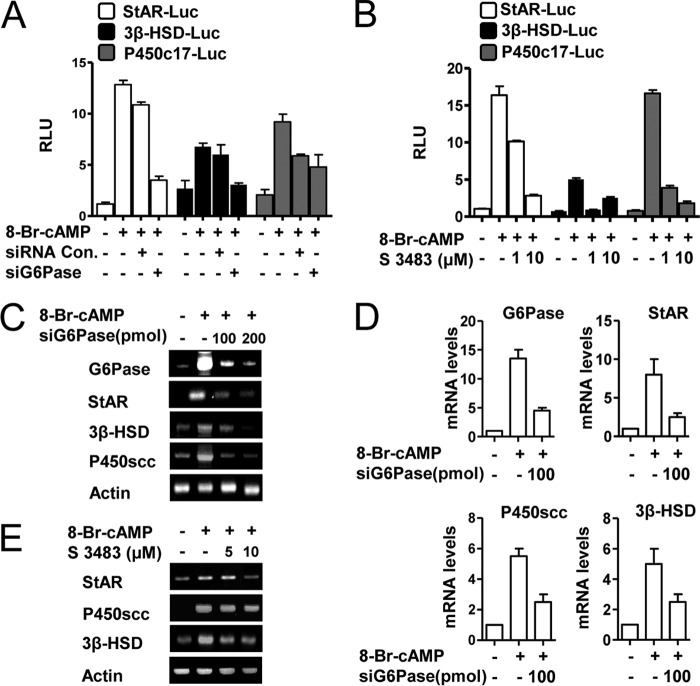
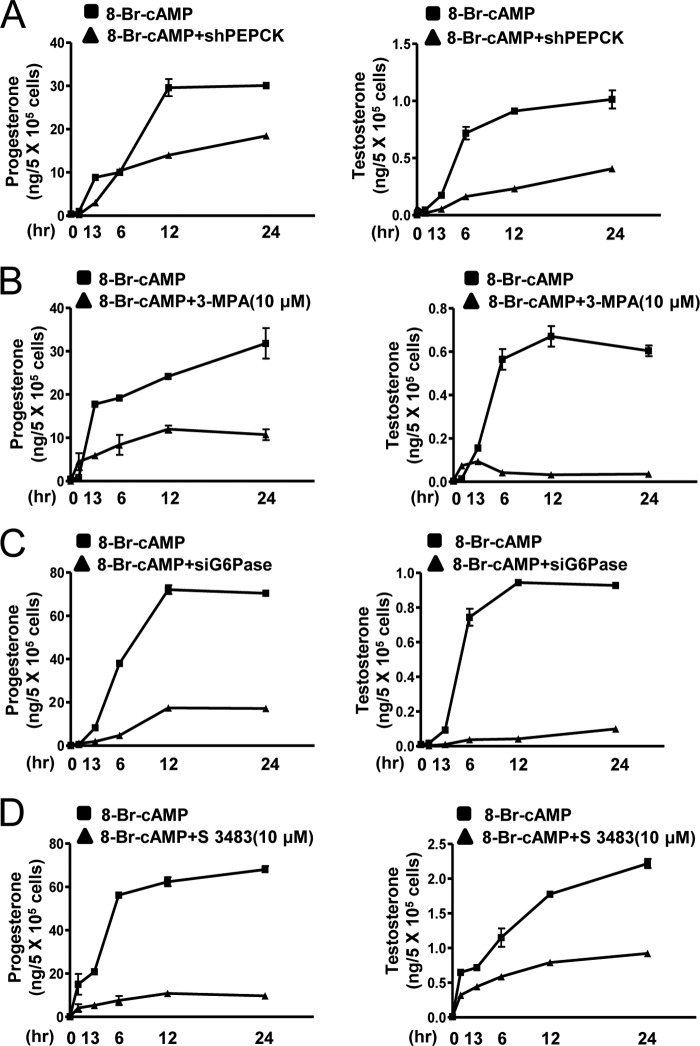
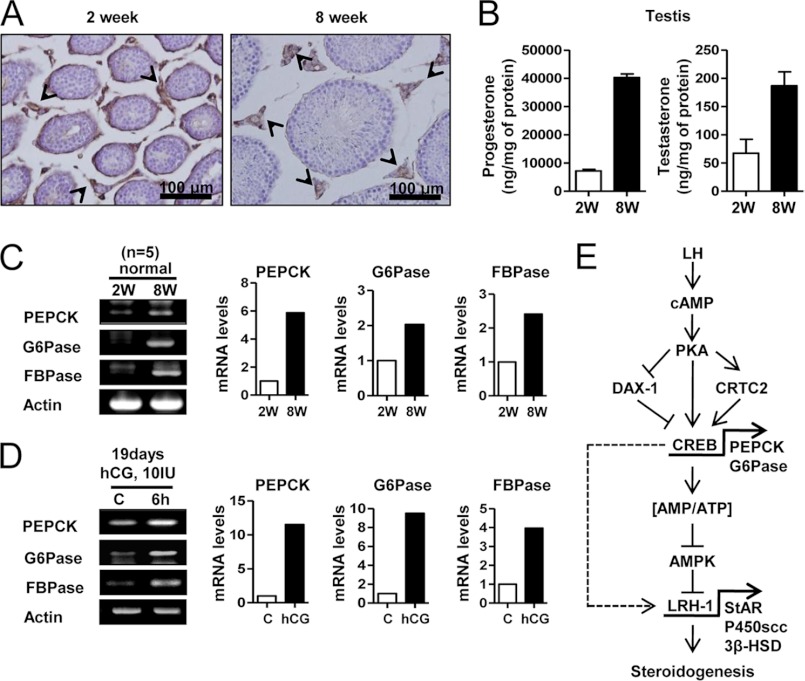
Cyclic AMP (cAMP) induces steroidogenic enzyme gene expression and stimulates testosterone production in Leydig cells. Phosphoenolpyruvate carboxykinase (PEPCK) is expressed in Leydig cells, but its role has not been defined. In this study, we found that PEPCK and glucose-6-phosphatase (Glc-6-Pase) are increased significantly following cAMP treatment of mouse Leydig cells. Moreover, cAMP treatment increased recruitment of the cAMP-response element-binding transcription factor and decreased recruitment of the corepressor DAX-1 on the pepck promoter. Furthermore, cAMP induced an increase in ATP that correlated with a decrease in phospho-AMP-activated protein kinase (AMPK). In contrast, knockdown or inhibition of PEPCK decreased ATP and increased phospho-AMPK. Treatment with an AMPK activator or overexpression of the constitutively active form of AMPK inhibited cAMP-induced steroidogenic enzyme promoter activities and gene expression. Liver receptor homolog-1 (LRH-1) was involved in cAMP-induced steroidogenic enzyme gene expression but was inhibited by AMPK activation in Leydig cells. Additionally, inhibition or knockdown of PEPCK and Glc-6-Pase decreased cAMP-mediated induction of steroidogenic enzyme gene expression and steroidogenesis. Finally, pubertal mouse (8-week-old) testes and human chorionic gonadotropin-induced prepubertal mouse testes showed increased PEPCK and Glc-6-Pase gene expression. Taken together, these results suggest that induction of PEPCK and Glc-6-Pase by cAMP plays an important role in Leydig cell steroidogenesis.
Figures
Similar articles
-
Insulin directly regulates steroidogenesis via induction of the orphan nuclear receptor DAX-1 in testicular Leydig cells.J Biol Chem. 2013 May 31;288(22):15937-46. doi: 10.1074/jbc.M113.451773. Epub 2013 Apr 15. J Biol Chem. 2013. PMID: 23589295 Free PMC article.
-
AMPK-dependent repression of hepatic gluconeogenesis via disruption of CREB.CRTC2 complex by orphan nuclear receptor small heterodimer partner.J Biol Chem. 2010 Oct 15;285(42):32182-91. doi: 10.1074/jbc.M110.134890. Epub 2010 Aug 5. J Biol Chem. 2010. PMID: 20688914 Free PMC article.
-
Sodium arsenite induces orphan nuclear receptor SHP gene expression via AMP-activated protein kinase to inhibit gluconeogenic enzyme gene expression.Am J Physiol Endocrinol Metab. 2008 Aug;295(2):E368-79. doi: 10.1152/ajpendo.00800.2007. Epub 2008 May 27. Am J Physiol Endocrinol Metab. 2008. PMID: 18505831
-
Hormonal regulation of steroidogenic enzyme gene expression in Leydig cells.J Steroid Biochem Mol Biol. 1992 Dec;43(8):895-906. doi: 10.1016/0960-0760(92)90317-C. J Steroid Biochem Mol Biol. 1992. PMID: 22217834 Review.
-
Transcription Factors in the Regulation of Leydig Cell Gene Expression and Function.Front Endocrinol (Lausanne). 2022 Apr 7;13:881309. doi: 10.3389/fendo.2022.881309. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35464056 Free PMC article. Review.
Cited by
-
Adiponectin influences progesterone production from MA-10 Leydig cells in a dose-dependent manner.Endocrine. 2015 Apr;48(3):957-67. doi: 10.1007/s12020-014-0456-y. Epub 2014 Oct 22. Endocrine. 2015. PMID: 25338202
-
PDK4 Deficiency Suppresses Hepatic Glucagon Signaling by Decreasing cAMP Levels.Diabetes. 2018 Oct;67(10):2054-2068. doi: 10.2337/db17-1529. Epub 2018 Jul 31. Diabetes. 2018. PMID: 30065033 Free PMC article.
-
Implications of adiponectin in linking metabolism to testicular function.Endocrine. 2014 May;46(1):16-28. doi: 10.1007/s12020-013-0102-0. Epub 2013 Nov 28. Endocrine. 2014. PMID: 24287788 Review.
-
Bone morphogenetic protein 4 inhibits rat stem/progenitor Leydig cell development and regeneration via SMAD-dependent and SMAD-independent signaling.Cell Death Dis. 2022 Dec 13;13(12):1039. doi: 10.1038/s41419-022-05471-8. Cell Death Dis. 2022. PMID: 36513649 Free PMC article.
-
Insulin directly regulates steroidogenesis via induction of the orphan nuclear receptor DAX-1 in testicular Leydig cells.J Biol Chem. 2013 May 31;288(22):15937-46. doi: 10.1074/jbc.M113.451773. Epub 2013 Apr 15. J Biol Chem. 2013. PMID: 23589295 Free PMC article.
References
-
- Yánez A. J., Nualart F., Droppelmann C., Bertinat R., Brito M., Concha I. I., Slebe J. C. (2003) Broad expression of fructose-1,6-bisphosphatase and phosphoenolpyruvate carboxykinase provide evidence for gluconeogenesis in human tissues other than liver and kidney. J. Cell. Physiol. 197, 189–197 - PubMed
-
- Hanson R. W., Reshef L. (2003) Glyceroneogenesis revisited. Biochimie 85, 1199–1205 - PubMed
-
- Pilkis S. J., Granner D. K. (1992) Molecular physiology of the regulation of hepatic gluconeogenesis and glycolysis. Annu. Rev. Physiol. 54, 885–909 - PubMed
-
- Hanson R. W., Reshef L. (1997) Regulation of phosphoenolpyruvate carboxykinase (GTP) gene expression. Annu. Rev. Biochem. 66, 581–611 - PubMed
-
- Shaywitz A. J., Greenberg M. E. (1999) CREB. A stimulus-induced transcription factor activated by a diverse array of extracellular signals. Annu. Rev. Biochem. 68, 821–861 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
