

Structural insights into the substrate specificity of the Rhodopseudomonas palustris protein acetyltransferase RpPat: identification of a loop critical for recognition by RpPat
- PMID: 23076154
- PMCID: PMC3510837
- DOI: 10.1074/jbc.M112.417360
Structural insights into the substrate specificity of the Rhodopseudomonas palustris protein acetyltransferase RpPat: identification of a loop critical for recognition by RpPat
Abstract
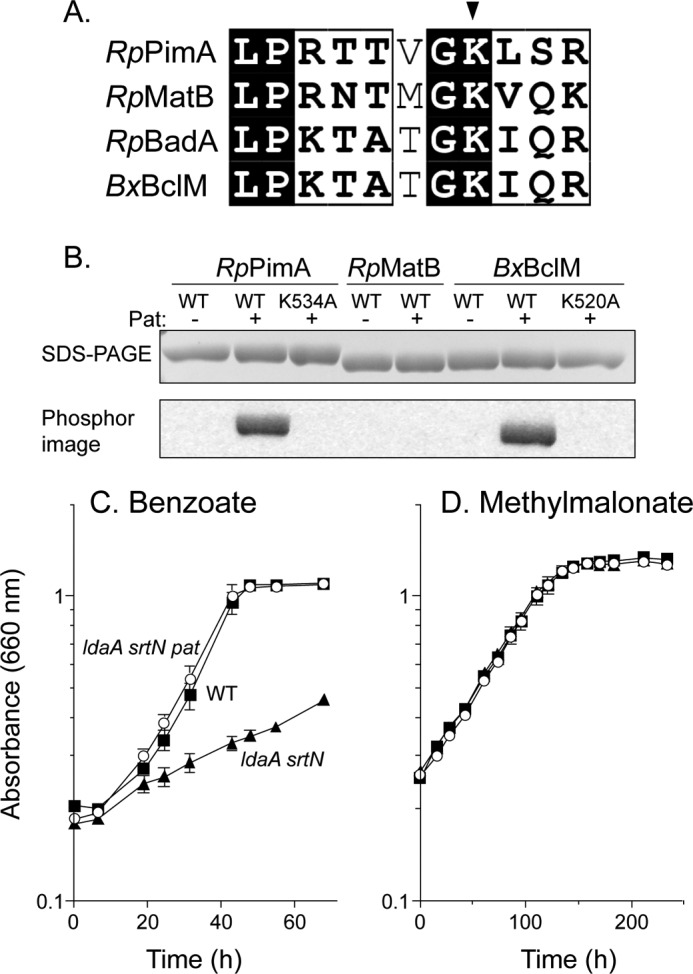
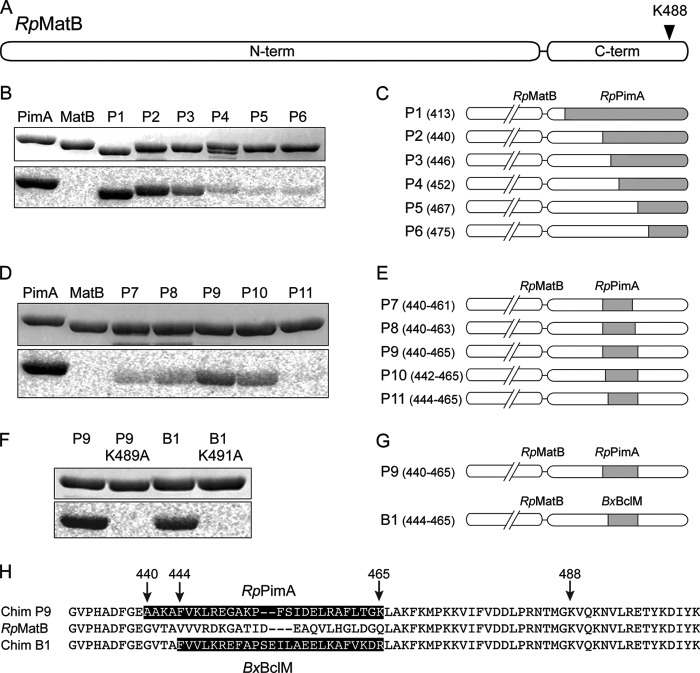
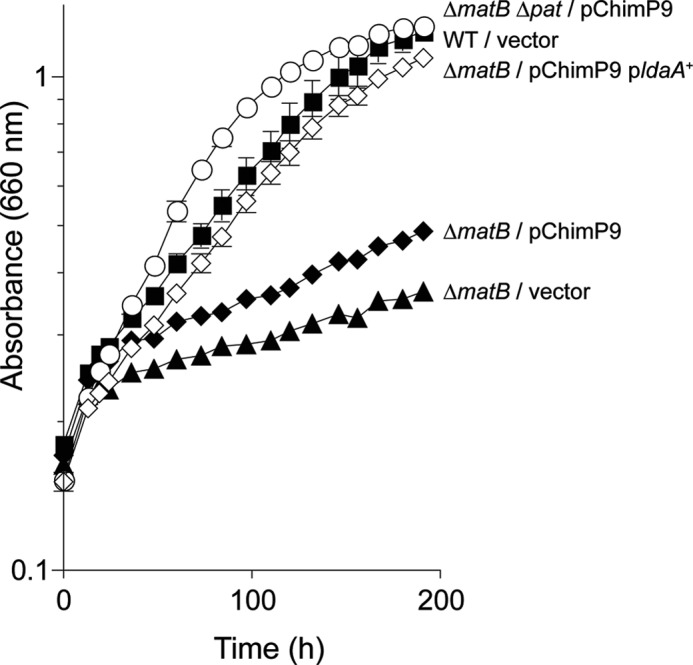
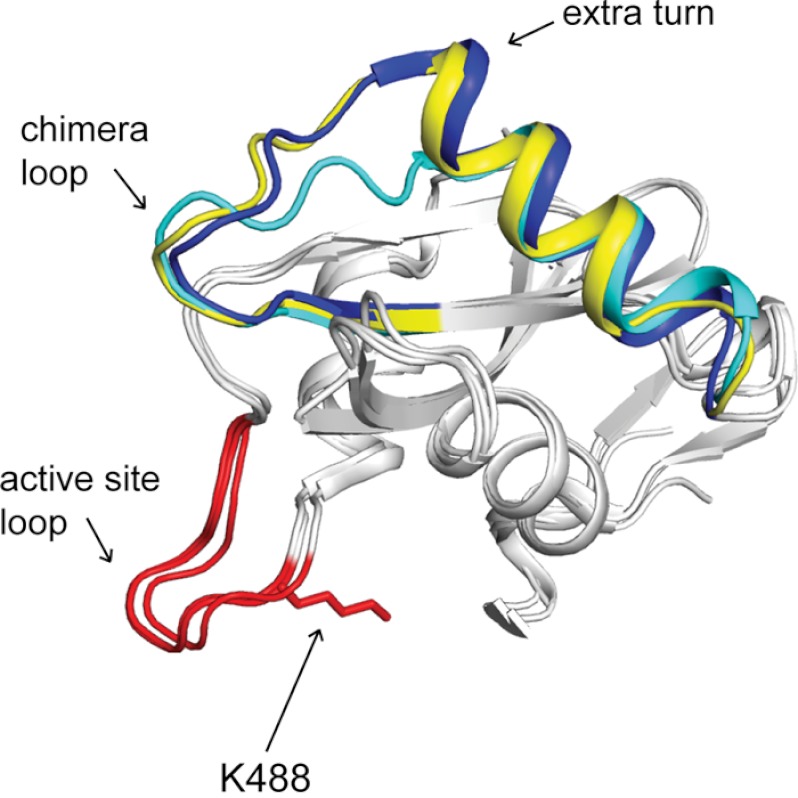
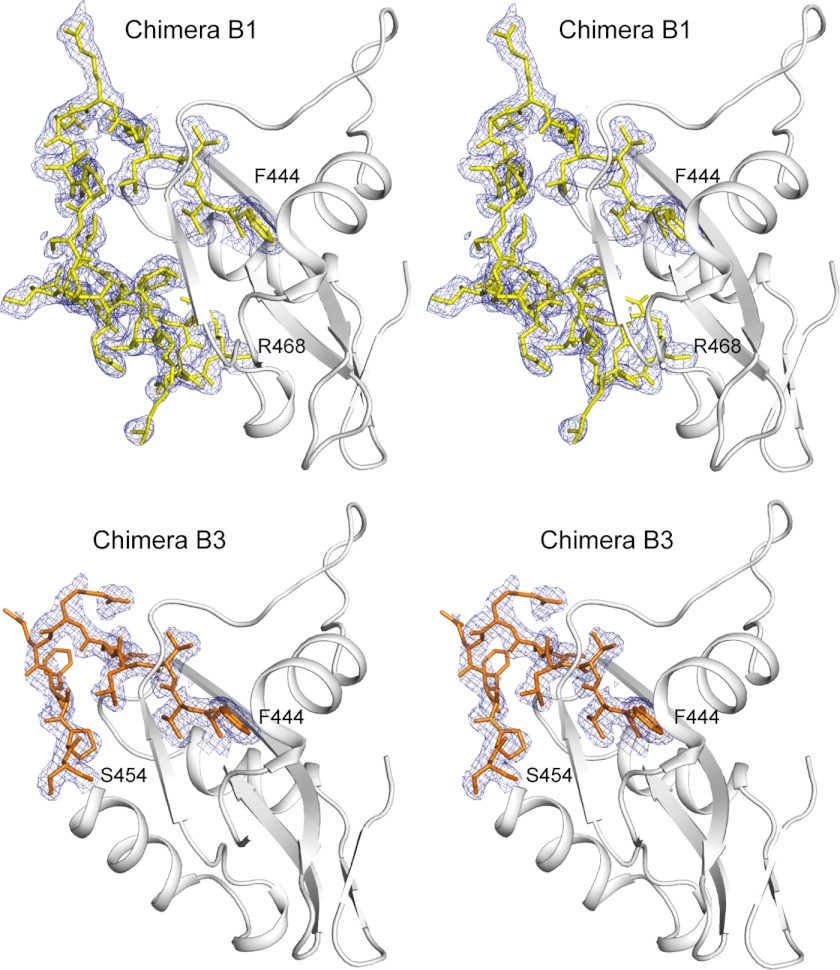
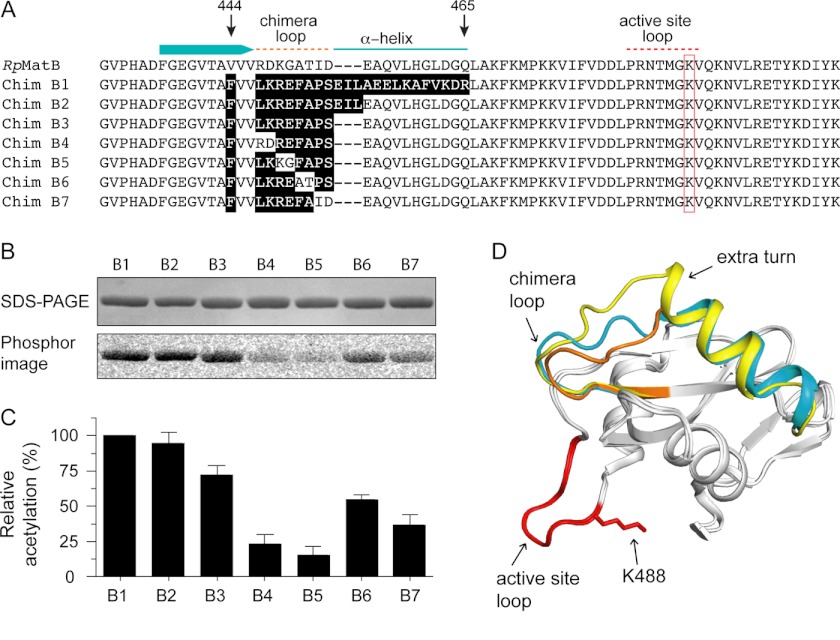
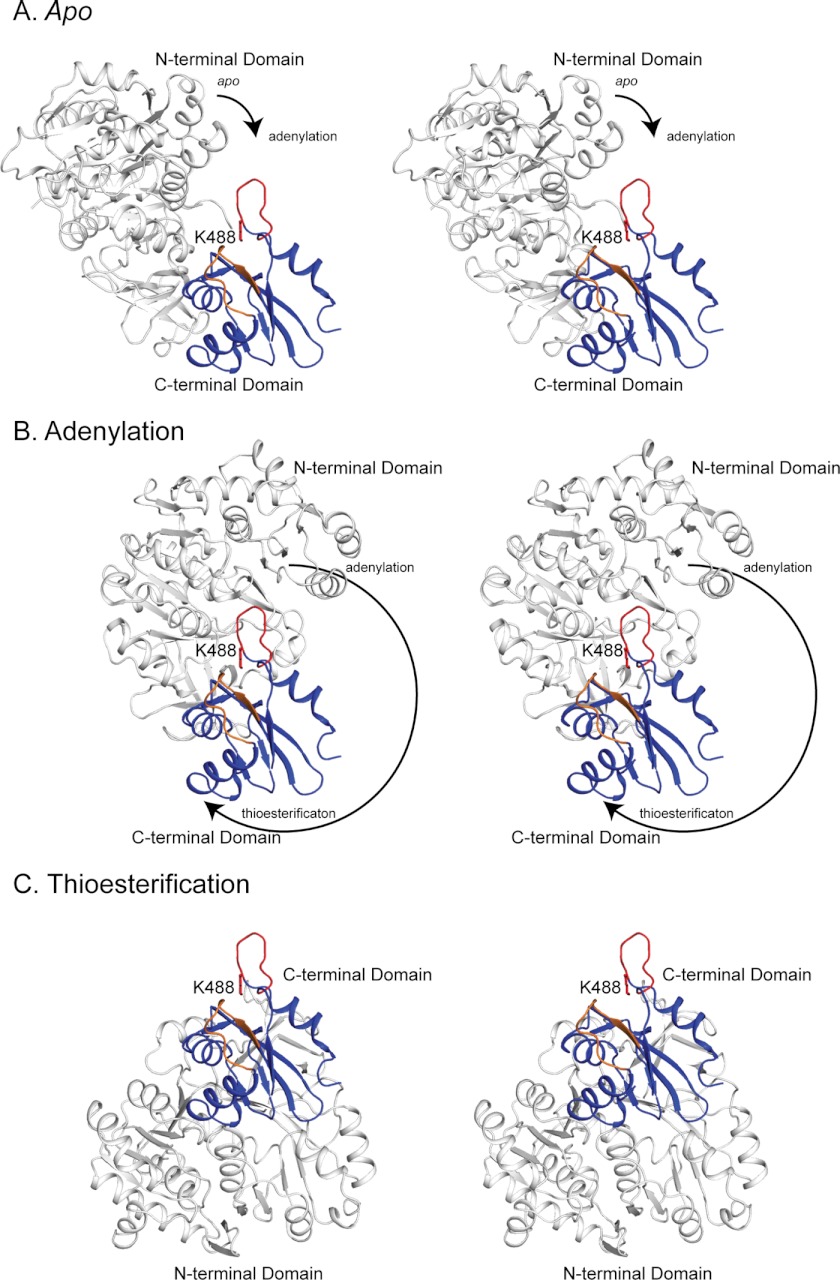
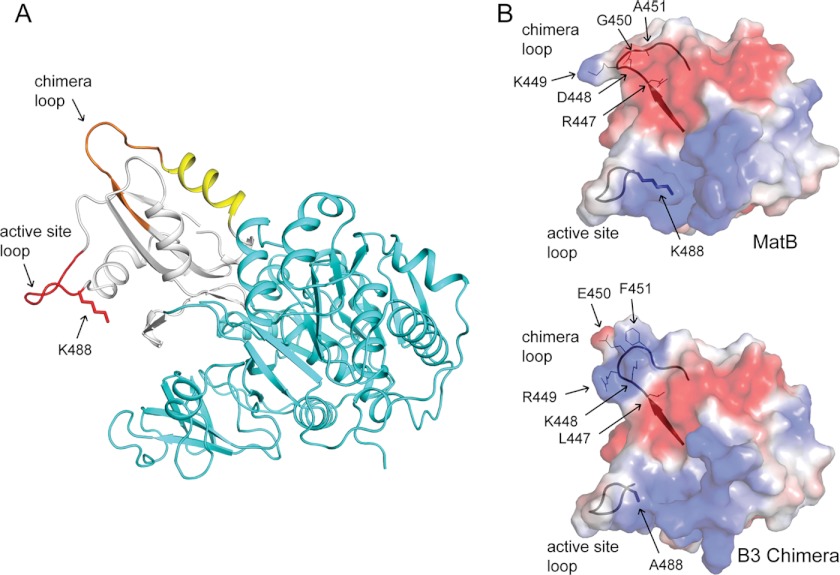
Lysine acetylation is a post-translational modification that is important for the regulation of metabolism in both prokaryotes and eukaryotes. In bacteria, the best studied protein acetyltransferase is Pat. In the purple photosynthetic bacterium Rhodopseudomonas palustris, at least 10 AMP-forming acyl-CoA synthetase enzymes are acetylated by the Pat homologue RpPat. All bona fide RpPat substrates contain the conserved motif PX(4)GK. Here, we show that the presence of such a motif is necessary but not sufficient for recognition by RpPat. RpPat failed to acetylate the methylmalonyl-CoA synthetase of this bacterium (hereafter RpMatB) in vivo and in vitro, despite the homology of RpMatB to known RpPat substrates. We used RpMatB to identify structural determinants that are recognized by RpPat. To do this, we constructed a series of RpMatB chimeras that became substrates of RpPat. In such chimeras, a short region (11-25 residues) of RpMatB located >20 residues N-terminal to the acetylation site was replaced with the corresponding sequences from other AMP-forming acyl-CoA synthetases that were known RpPat substrates. Strikingly, the enzymatic activity of RpMatB chimeras was regulated by acetylation both in vitro and in vivo. Crystal structures of two of these chimeras showed that the major difference between them and wild-type RpMatB was within a loop region ∼23 Å from the acetylation site. On the basis of these results, we suggest that RpPat likely interacts with a relatively large surface of its substrates, in addition to the PX(4)GK motif, and that RpPat probably has relatively narrow substrate specificity.
Figures
References
-
- Kim G. W., Yang X. J. (2011) Comprehensive lysine acetylomes emerging from bacteria to humans. Trends. Biochem. Sci. 36, 211–220 - PubMed
-
- Oberdoerffer P., Michan S., McVay M., Mostoslavsky R., Vann J., Park S. K., Hartlerode A., Stegmuller J., Hafner A., Loerch P., Wright S. M., Mills K. D., Bonni A., Yankner B. A., Scully R., Prolla T. A., Alt F. W., Sinclair D. A. (2008) SIRT1 redistribution on chromatin promotes genomic stability but alters gene expression during aging. Cell 135, 907–918 - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
