

The unsilent majority-TRPV1 drives "spontaneous" transmission of unmyelinated primary afferents within cardiorespiratory NTS
- PMID: 23076872
- PMCID: PMC3532589
- DOI: 10.1152/ajpregu.00398.2012
The unsilent majority-TRPV1 drives "spontaneous" transmission of unmyelinated primary afferents within cardiorespiratory NTS
Abstract
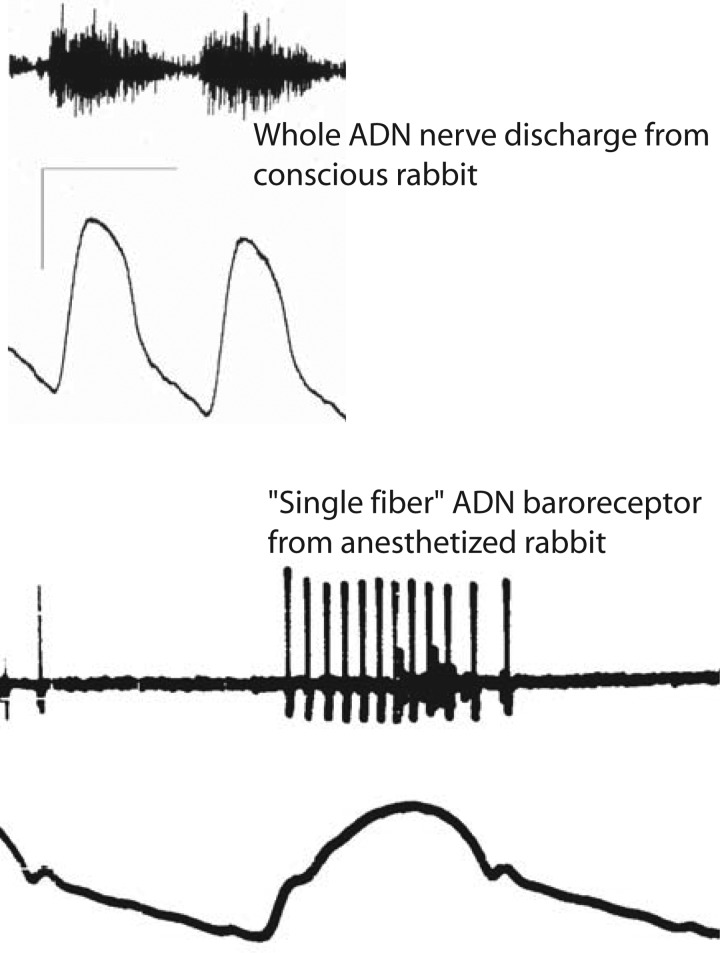
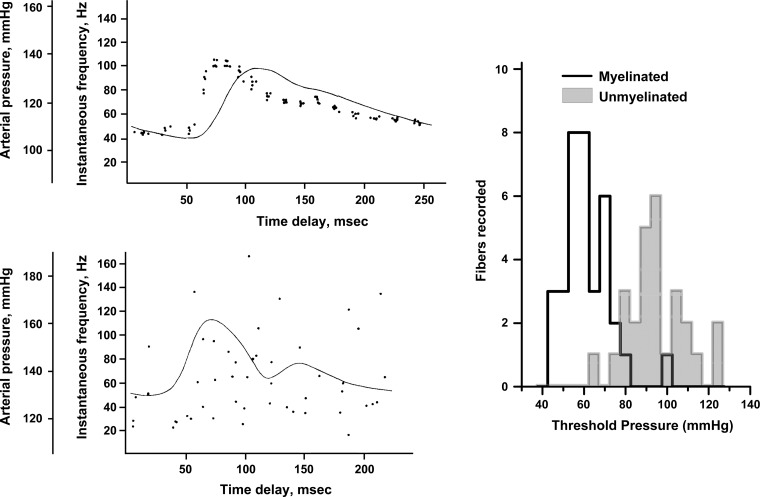
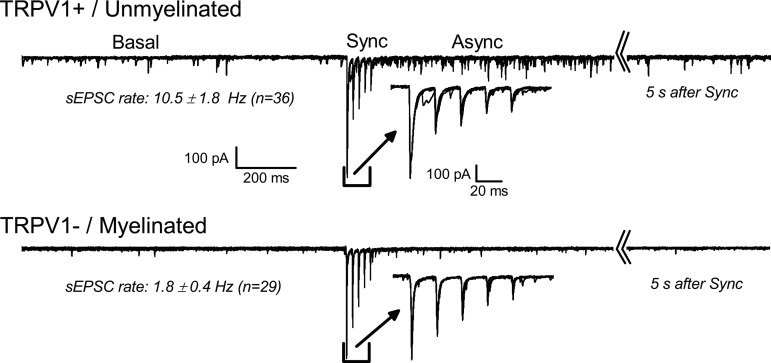
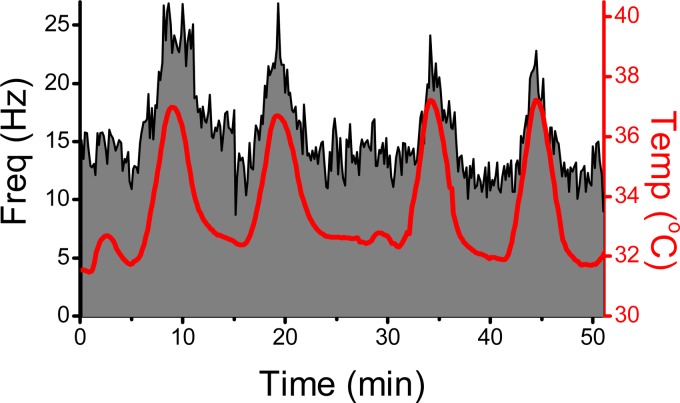
Cranial primary afferent sensory neurons figure importantly in homeostatic control of visceral organ systems. Of the two broad classes of visceral afferents, the role of unmyelinated or C-type class remains poorly understood. This review contrasts key aspects of peripheral discharge properties of C-fiber afferents and their glutamate transmission mechanisms within the solitary tract nucleus (NTS). During normal prevailing conditions, most information arrives at the NTS through myelinated A-type nerves. However, most of visceral afferent axons (75-90%) in NTS are unmyelinated, C-type axons. Centrally, C-type solitary tract (ST) afferent terminals have presynaptic transient receptor potential vanilloid type 1 (TRPV1) receptors. Capsaicin activation of TRPV1 blocks phasic or synchronous release of glutamate but facilitates release of glutamate from a separate pool of vesicles. This TRPV1-operated pool of vesicles is active at normal temperatures and is responsible for actively driving a 10-fold higher release of glutamate at TRPV1 compared with TRPV1- terminals even in the absence of afferent action potentials. This novel TRPV1 mechanism is responsible for an additional asynchronous release of glutamate that is not present in myelinated terminals. The NTS is rich with presynaptic G protein-coupled receptors, and the implications of TRPV1-operated glutamate offer unique targets for signaling in C-type sensory afferent terminals from neuropeptides, inflammatory mediators, lipid metabolites, cytokines, and cannabinoids. From a homeostatic view, this combination could have broad implications for integration in chronic pathological disturbances in which the numeric dominance of C-type endings and TRPV1 would broadly disturb multisystem control mechanisms.
Figures
References
-
- Adrian ED. Nobel Lecture: The activity of the nerve fibres (1932). In: Nobel Lectures: Physiology or Medicine 1922–1941 Amsterdam: Elsevier, 1965
-
- Agarwal SK, Calaresu FR. Reciprocal connections between nucleus tractus solitarii and rostral ventrolateral medulla. Brain Res 523: 305–308, 1990 - PubMed
-
- Akopian AN, Abson NC, Wood JN. Molecular genetic approaches to nociceptor development and function. Trends Neurosci 19: 240–246, 1996 - PubMed
-
- Akopian AN, Sivilotti L, Wood JN. A tetrodotoxin-resistant voltage-gated sodium channel expressed by sensory neurons. Nature 379: 257–262, 1996 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources