

Investigation of influenza virus polymerase activity in pig cells
- PMID: 23077313
- PMCID: PMC3536367
- DOI: 10.1128/JVI.01633-12
Investigation of influenza virus polymerase activity in pig cells
Abstract
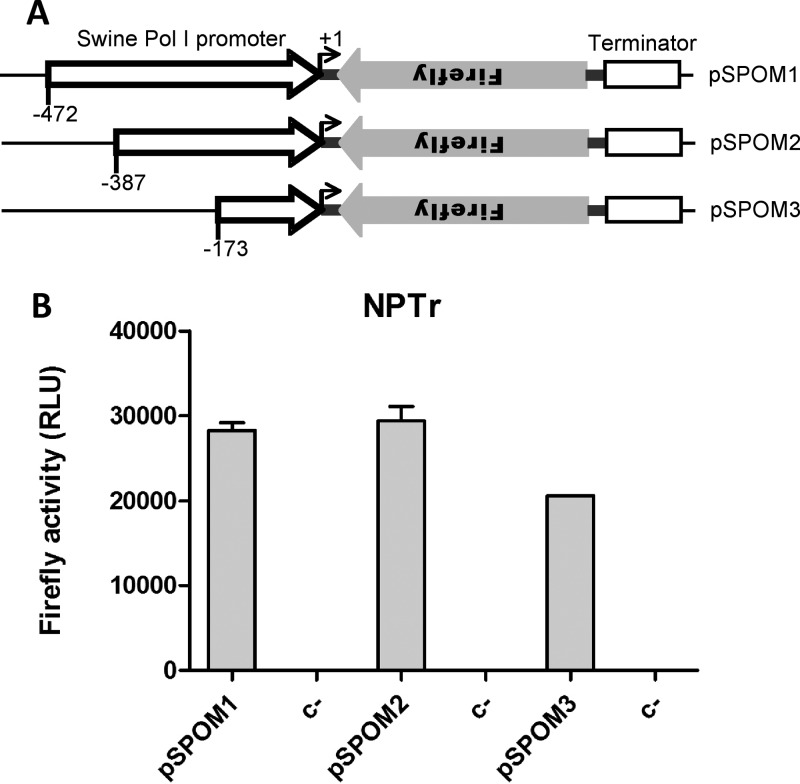
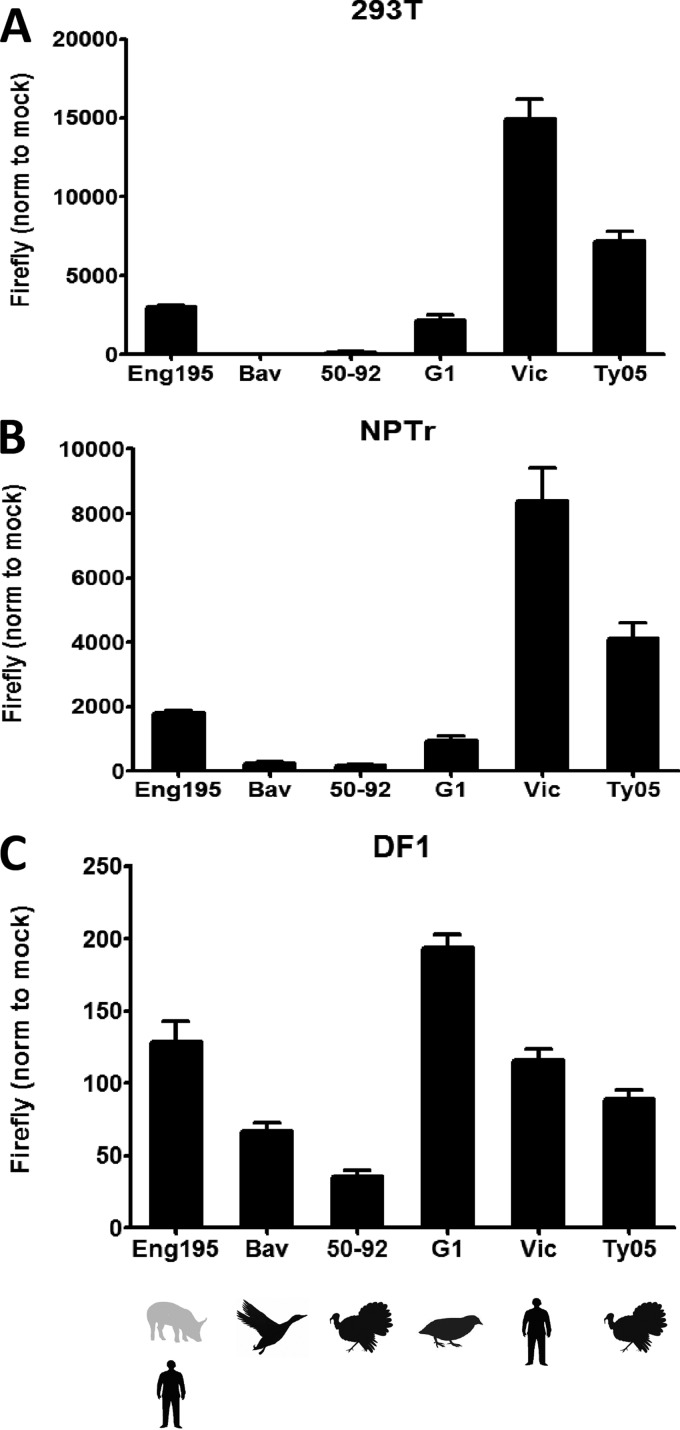
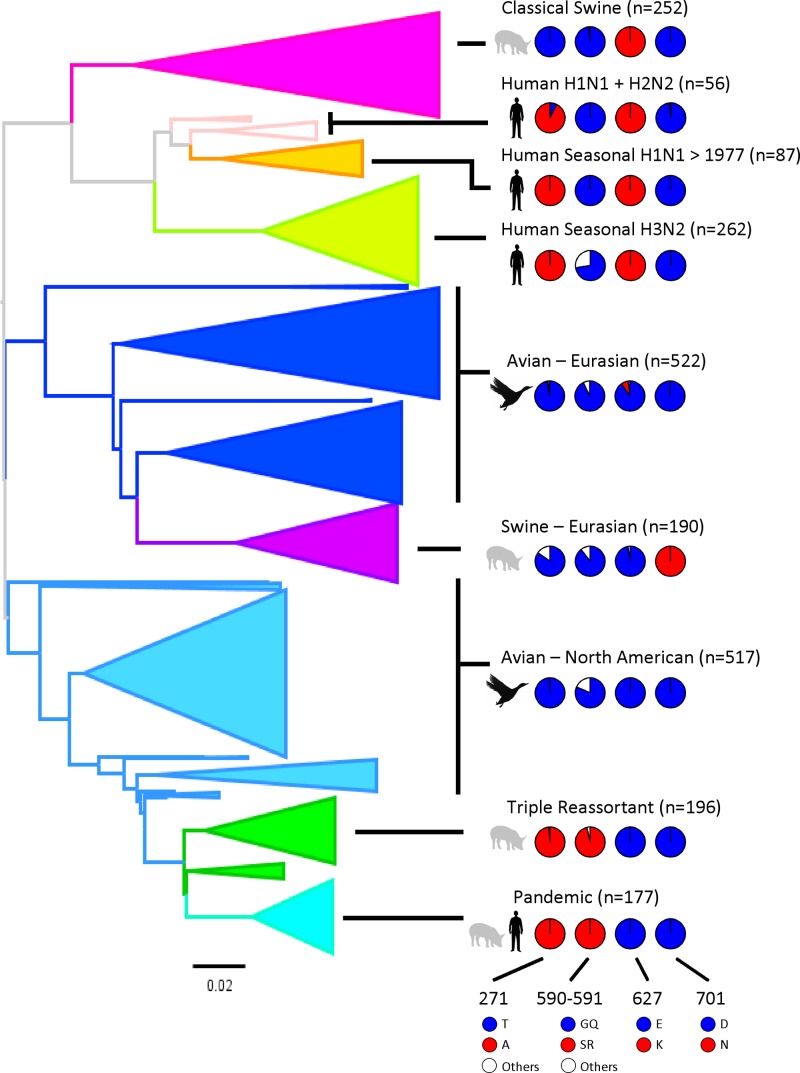
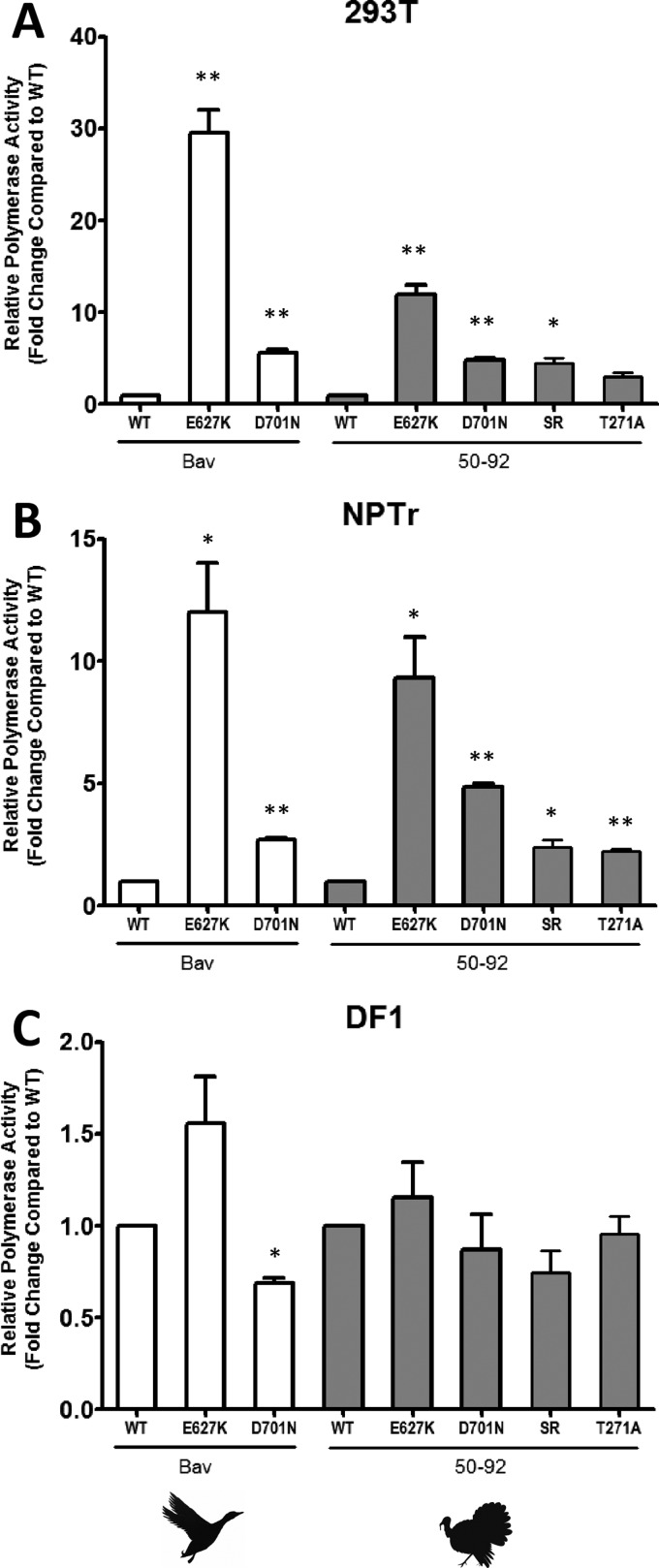
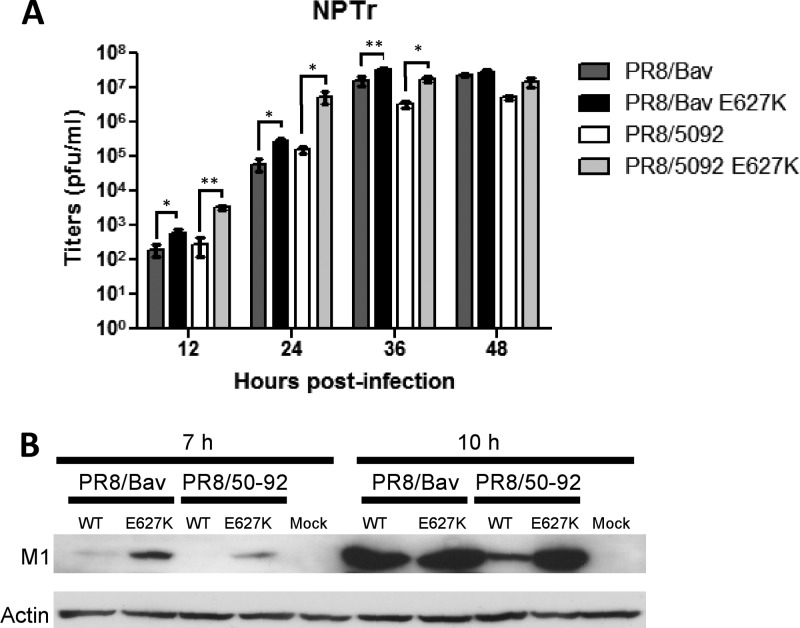
Reassortant influenza viruses with combinations of avian, human, and/or swine genomic segments have been detected frequently in pigs. As a consequence, pigs have been accused of being a "mixing vessel" for influenza viruses. This implies that pig cells support transcription and replication of avian influenza viruses, in contrast to human cells, in which most avian influenza virus polymerases display limited activity. Although influenza virus polymerase activity has been studied in human and avian cells for many years by use of a minigenome assay, similar investigations in pig cells have not been reported. We developed the first minigenome assay for pig cells and compared the activities of polymerases of avian or human influenza virus origin in pig, human, and avian cells. We also investigated in pig cells the consequences of some known mammalian host range determinants that enhance influenza virus polymerase activity in human cells, such as PB2 mutations E627K, D701N, G590S/Q591R, and T271A. The two typical avian influenza virus polymerases used in this study were poorly active in pig cells, similar to what is seen in human cells, and mutations that adapt the avian influenza virus polymerase for human cells also increased activity in pig cells. In contrast, a different pattern was observed in avian cells. Finally, highly pathogenic avian influenza virus H5N1 polymerase activity was tested because this subtype has been reported to replicate only poorly in pigs. H5N1 polymerase was active in swine cells, suggesting that other barriers restrict these viruses from becoming endemic in pigs.
Figures
References
-
- Alexander DJ. 2000. A review of avian influenza in different bird species. Vet. Microbiol. 74:3–13 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
