

Metabolic signatures uncover distinct targets in molecular subsets of diffuse large B cell lymphoma
- PMID: 23079663
- PMCID: PMC3479446
- DOI: 10.1016/j.ccr.2012.08.014
Metabolic signatures uncover distinct targets in molecular subsets of diffuse large B cell lymphoma
Abstract
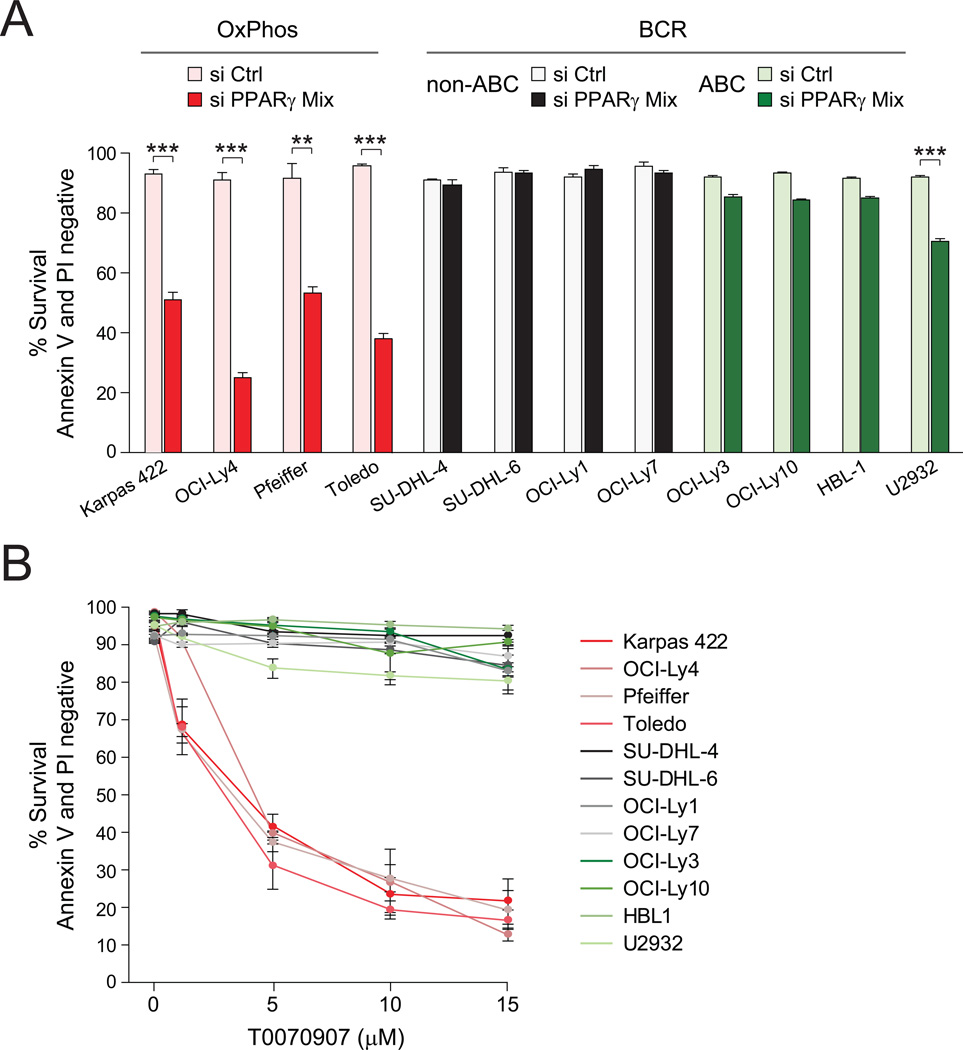
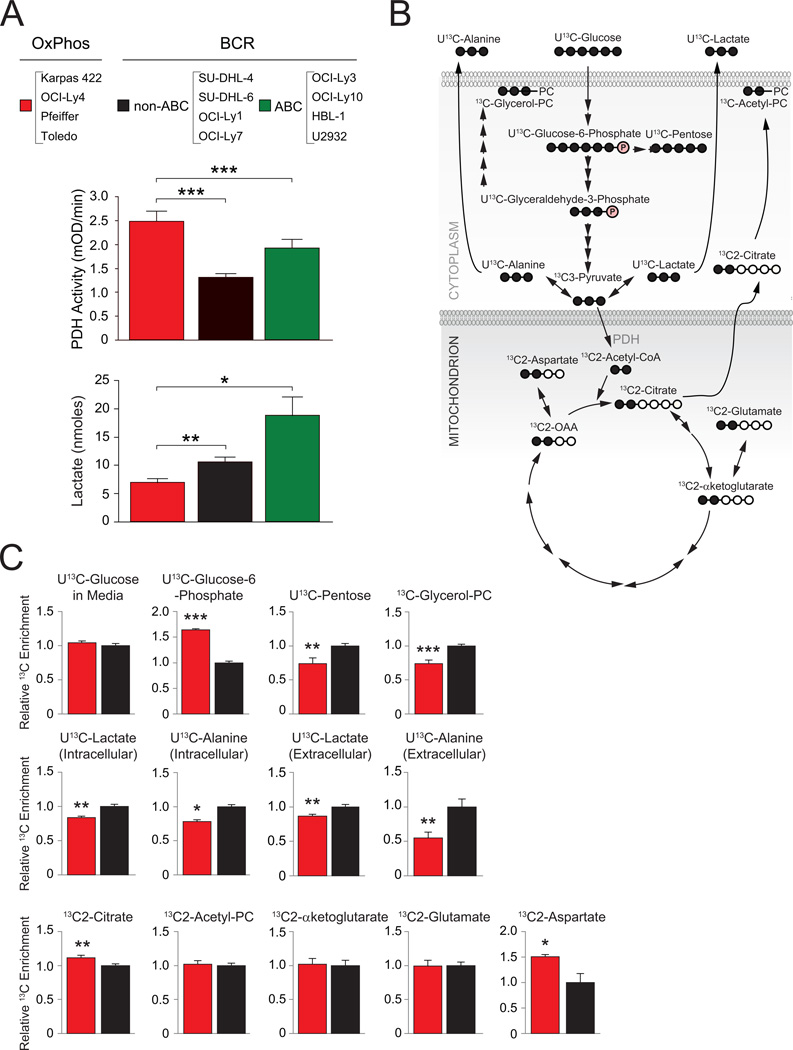
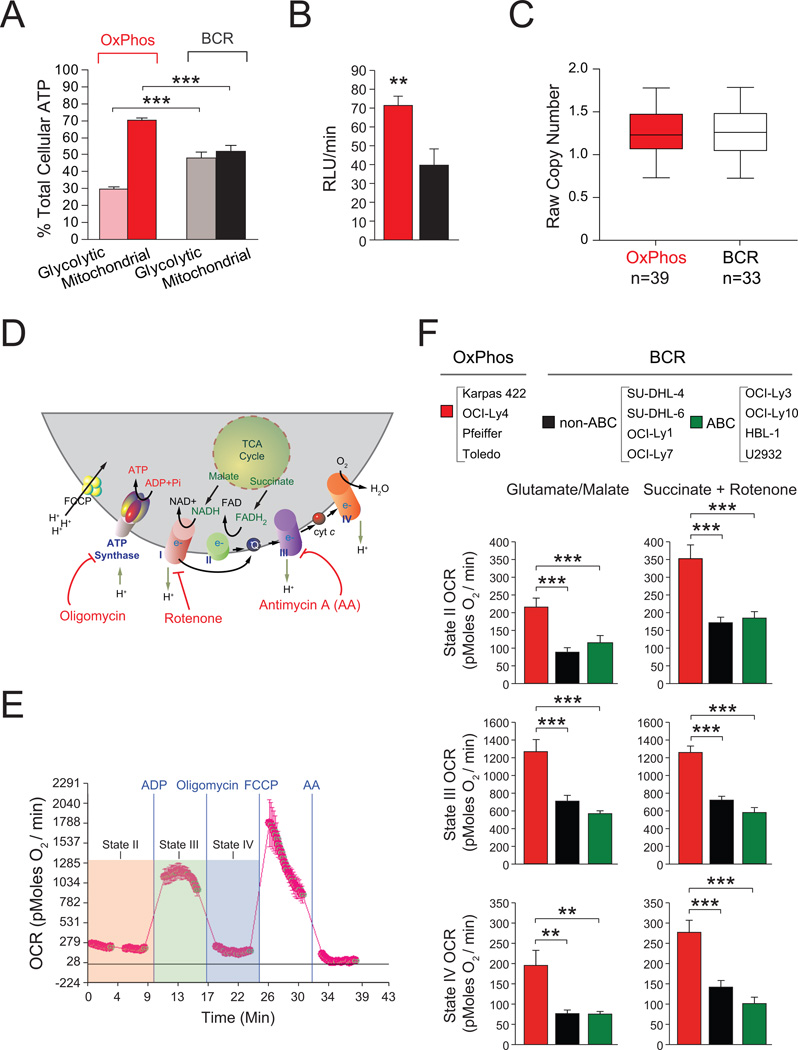
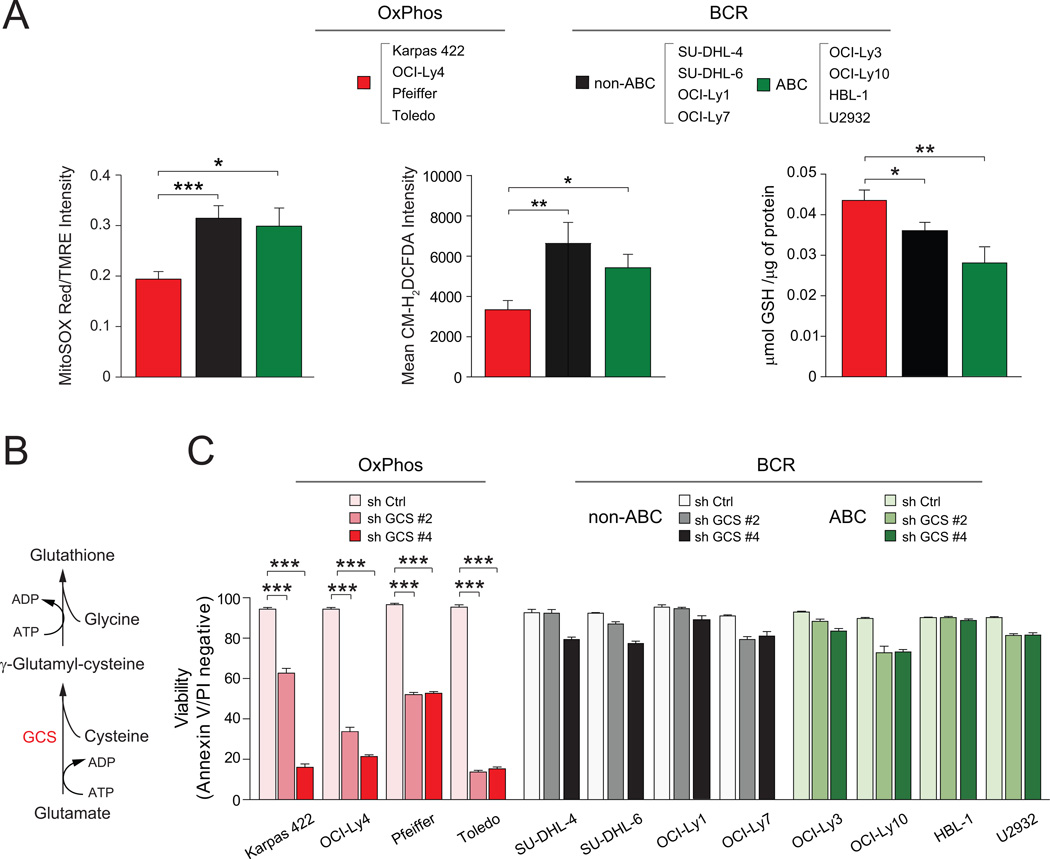
Molecular signatures have identified several subsets of diffuse large B cell lymphoma (DLBCL) and rational targets within the B cell receptor (BCR) signaling axis. The OxPhos-DLBCL subset, which harbors the signature of genes involved in mitochondrial metabolism, is insensitive to inhibition of BCR survival signaling but is functionally undefined. We show that, compared with BCR-DLBCLs, OxPhos-DLBCLs display enhanced mitochondrial energy transduction, greater incorporation of nutrient-derived carbons into the tricarboxylic acid cycle, and increased glutathione levels. Moreover, perturbation of the fatty acid oxidation program and glutathione synthesis proved selectively toxic to this tumor subset. Our analysis provides evidence for distinct metabolic fingerprints and associated survival mechanisms in DLBCL and may have therapeutic implications.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
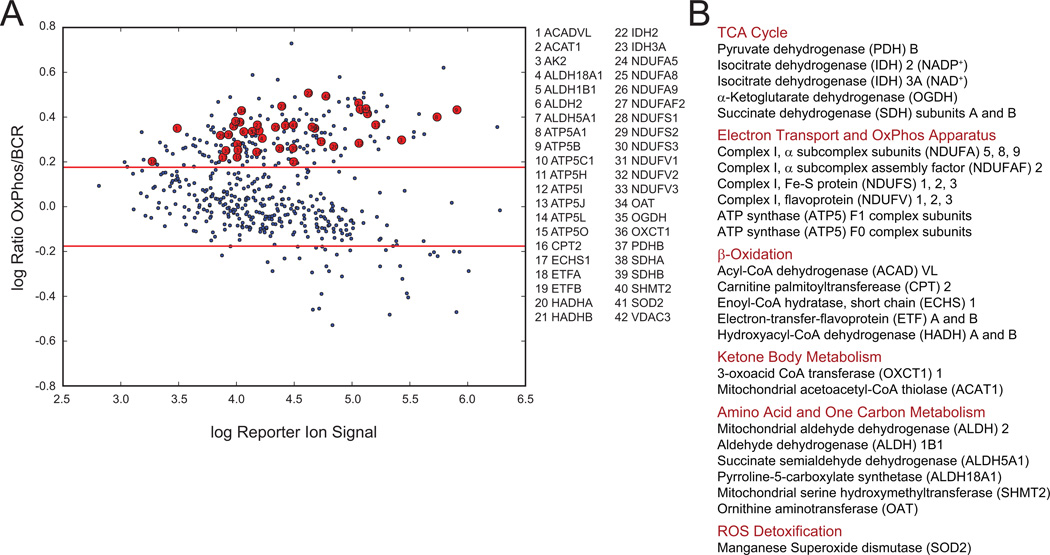
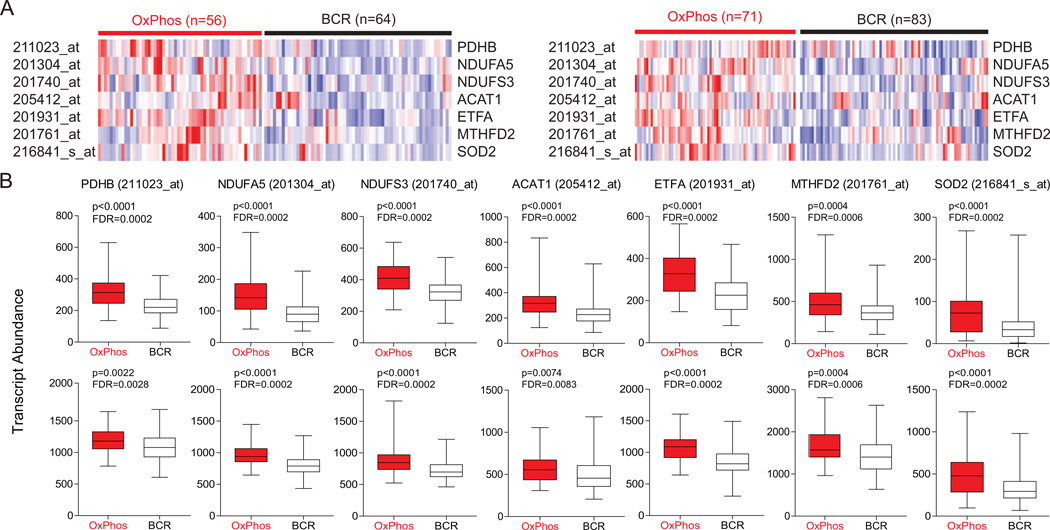
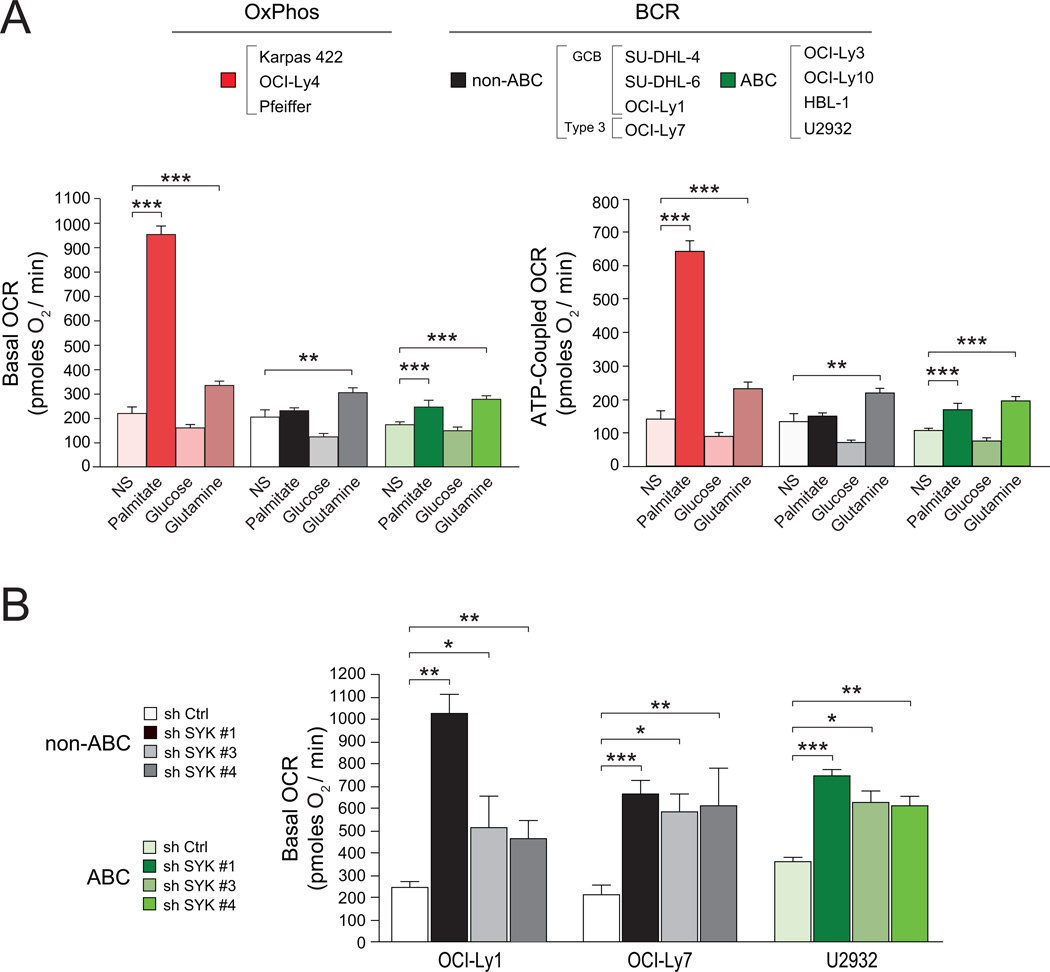
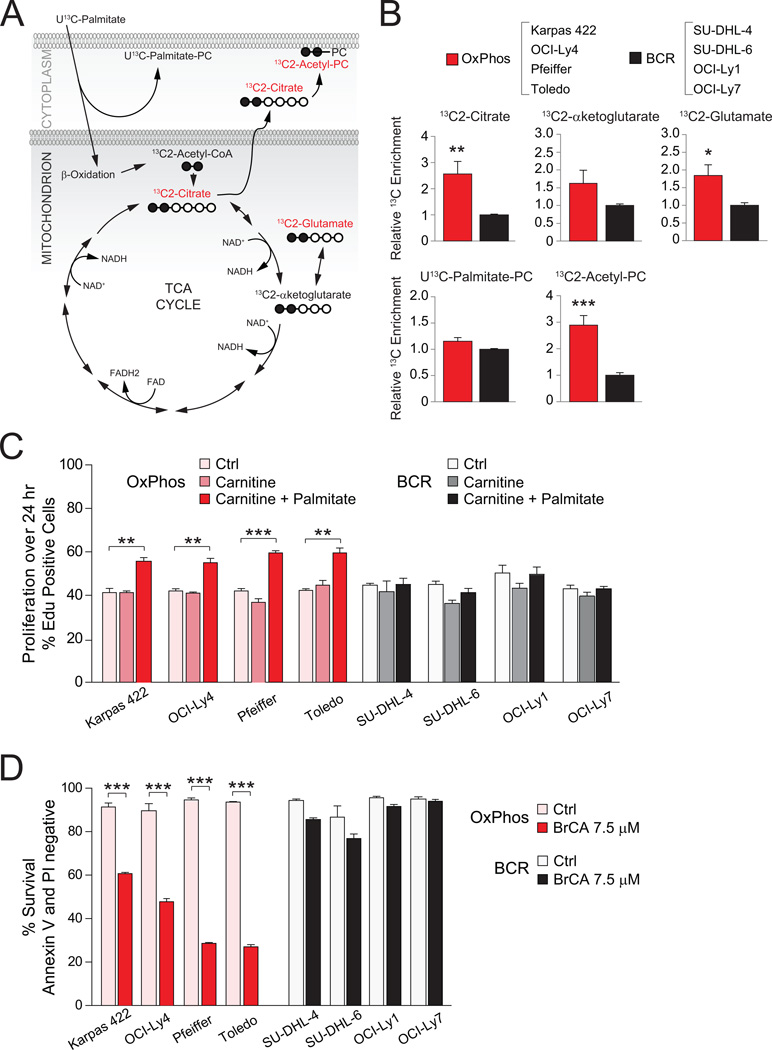
Comment in
-
A mitochondrial power play in lymphoma.Cancer Cell. 2012 Oct 16;22(4):423-4. doi: 10.1016/j.ccr.2012.09.023. Cancer Cell. 2012. PMID: 23079653 Free PMC article.
References
-
- Abramson JS, Shipp MA. Advances in the biology and therapy of diffuse large B-cell lymphoma: moving toward a molecularly targeted approach. Blood. 2005;106:1164–1174. - PubMed
-
- Alizadeh AA, Eisen MB, Davis RE, Ma C, Lossos IS, Rosenwald A, Boldrick JC, Sabet H, Tran T, Yu X, et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature. 2000;403:503–511. - PubMed
-
- Barger JF, Plas DR. Balancing biosynthesis and bioenergetics: metabolic programs in oncogenesis. Endocr Relat Cancer. 2010;17:R287–R304. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
