

Slit/Robo signaling modulates the proliferation of central nervous system progenitors
- PMID: 23083737
- PMCID: PMC4443924
- DOI: 10.1016/j.neuron.2012.08.003
Slit/Robo signaling modulates the proliferation of central nervous system progenitors
Abstract
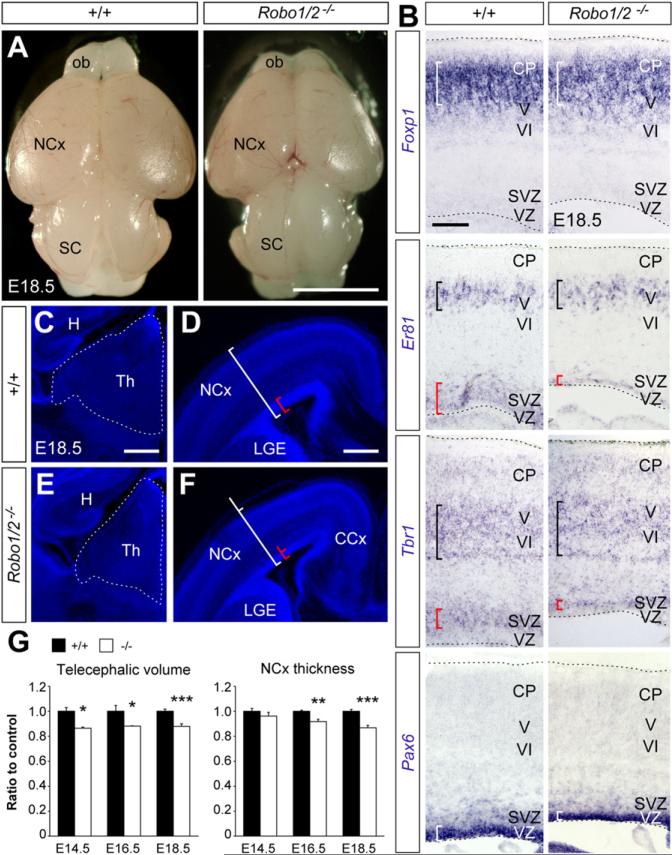
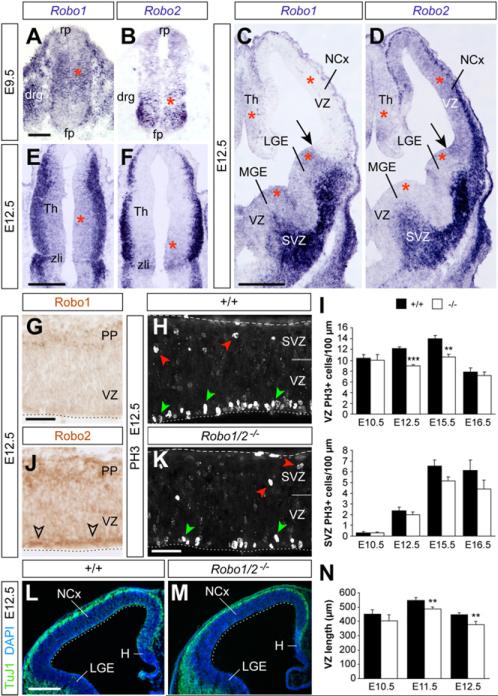
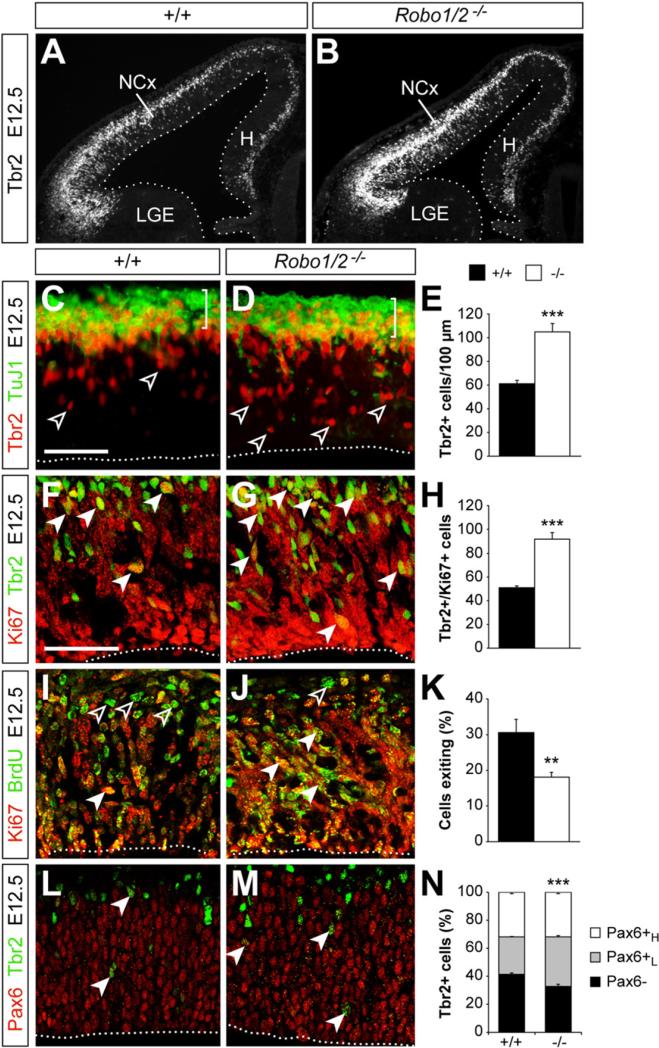
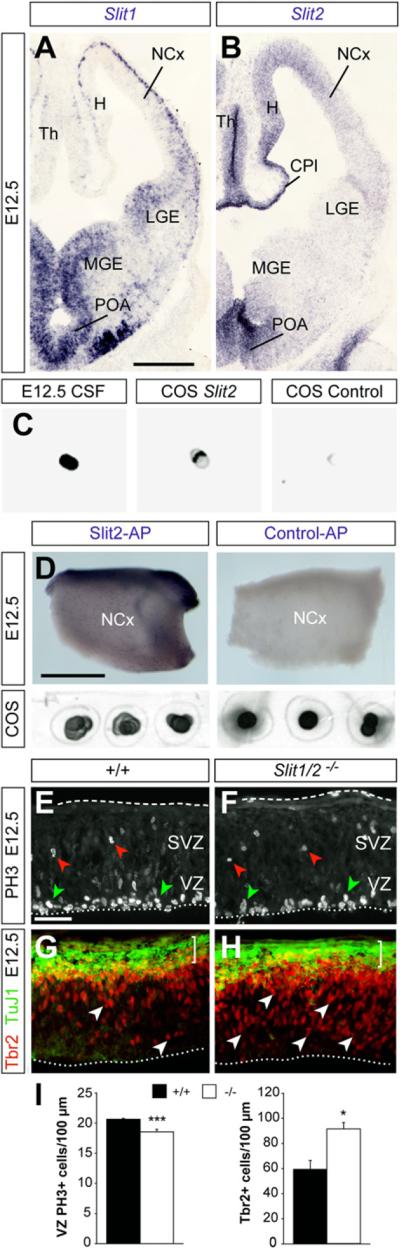
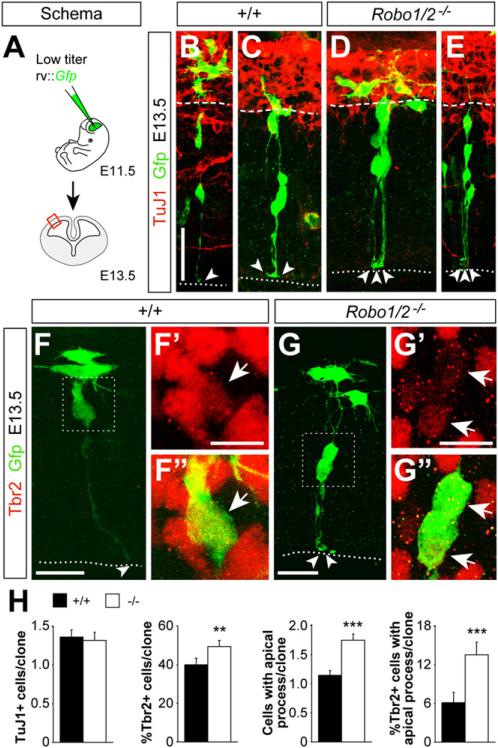
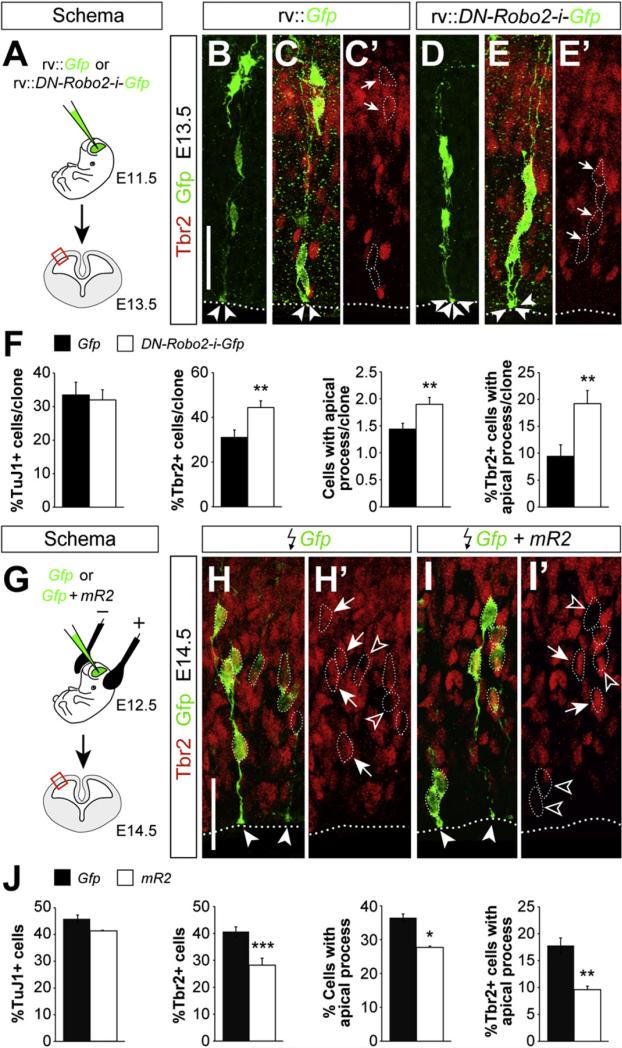
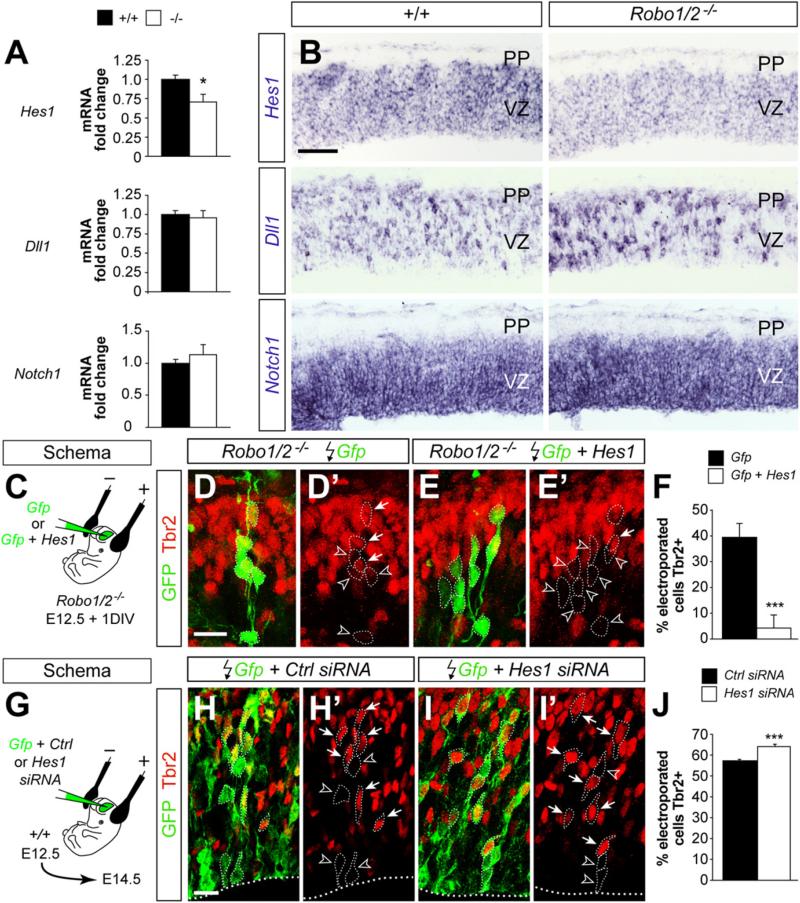
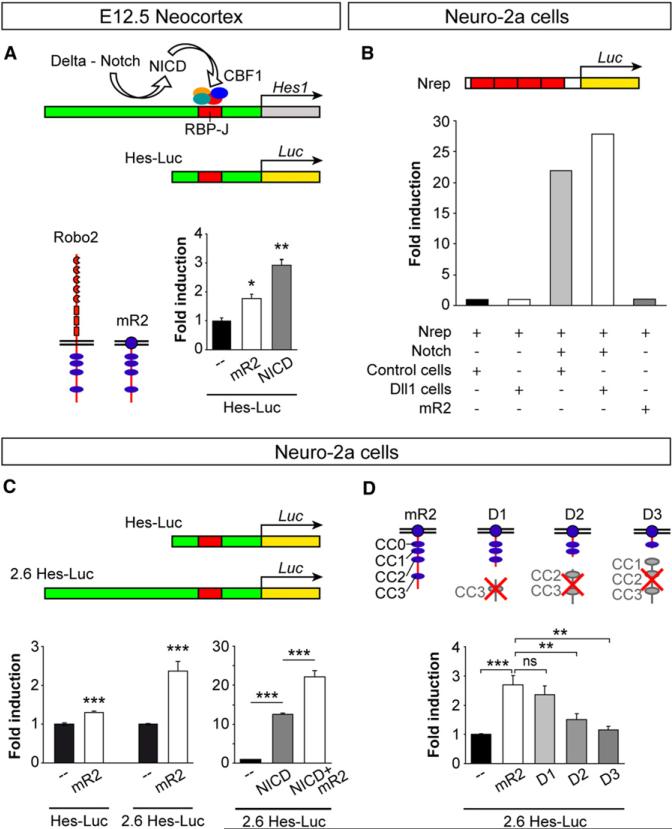
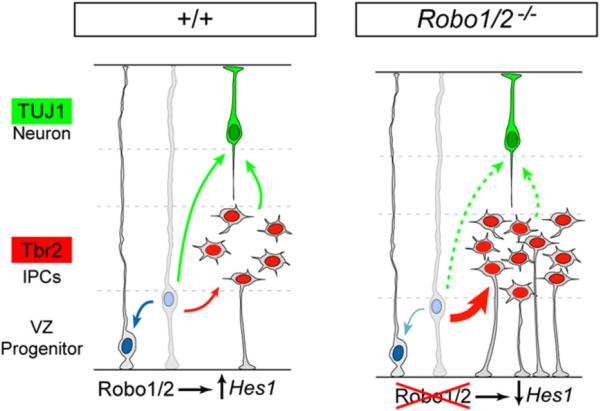
Neurogenesis relies on a delicate balance between progenitor maintenance and neuronal production. Progenitors divide symmetrically to increase the pool of dividing cells. Subsequently, they divide asymmetrically to self-renew and produce new neurons or, in some brain regions, intermediate progenitor cells (IPCs). Here we report that central nervous system progenitors express Robo1 and Robo2, receptors for Slit proteins that regulate axon guidance, and that absence of these receptors or their ligands leads to loss of ventricular mitoses. Conversely, production of IPCs is enhanced in Robo1/2 and Slit1/2 mutants, suggesting that Slit/Robo signaling modulates the transition between primary and intermediate progenitors. Unexpectedly, these defects do not lead to transient overproduction of neurons, probably because supernumerary IPCs fail to detach from the ventricular lining and cycle very slowly. At the molecular level, the role of Slit/Robo in progenitor cells involves transcriptional activation of the Notch effector Hes1. These findings demonstrate that Robo signaling modulates progenitor cell dynamics in the developing brain.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Andrews W, Barber M, Hernadez-Miranda LR, Xian J, Rakić S, Sundaresan V, Rabbitts TH, Pannell R, Rabbitts P, Thompson H, et al. The role of Slit-Robo signaling in the generation, migration and morphological differentiation of cortical interneurons. Dev. Biol. 2008;313:648–658. - PubMed
-
- Bagri A, Marín O, Plump AS, Mak J, Pleasure SJ, Rubenstein JL, Tessier-Lavigne M. Slit proteins prevent midline crossing and determine the dorsoventral position of major axonal pathways in the mammalian forebrain. Neuron. 2002;33:233–248. - PubMed
-
- Bashaw GJ, Kidd T, Murray D, Pawson T, Goodman CS. Repulsive axon guidance: Abelson and Enabled play opposing roles down-stream of the roundabout receptor. Cell. 2000;101:703–715. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
