

Calcium feedback to cGMP synthesis strongly attenuates single-photon responses driven by long rhodopsin lifetimes
- PMID: 23083739
- PMCID: PMC3594095
- DOI: 10.1016/j.neuron.2012.07.029
Calcium feedback to cGMP synthesis strongly attenuates single-photon responses driven by long rhodopsin lifetimes
Erratum in
- Neuron. 2013 Jun 5;78(5):949
Abstract
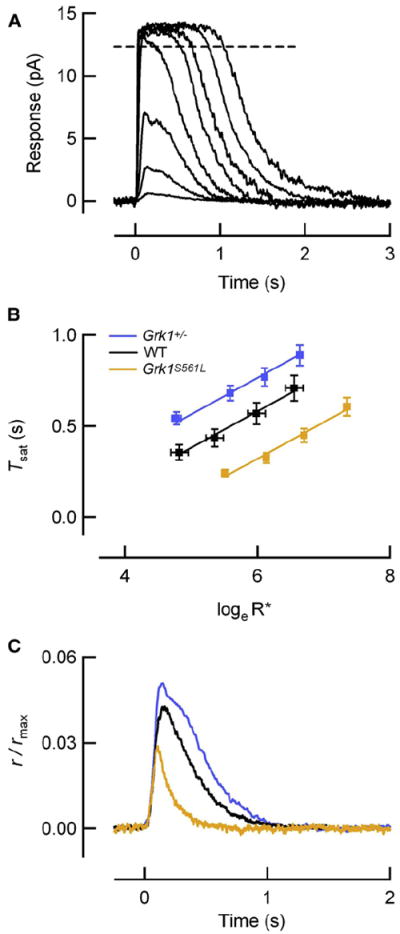
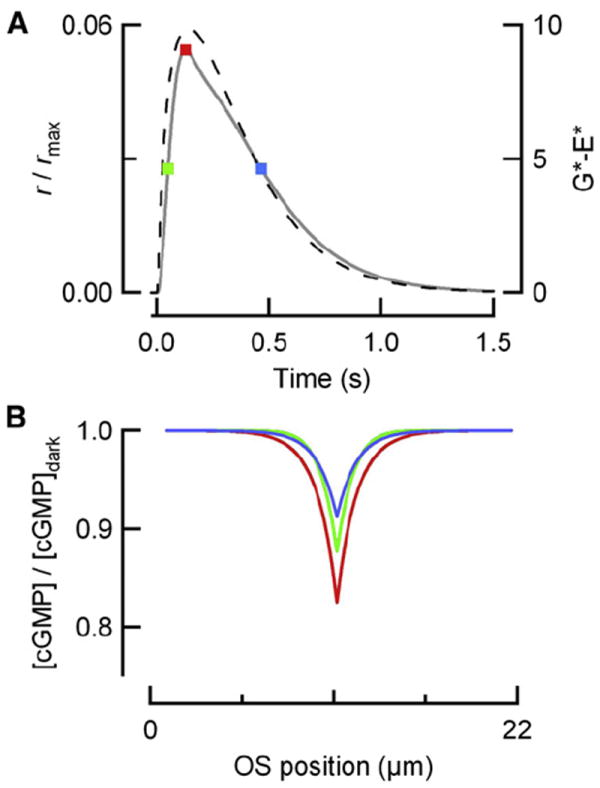
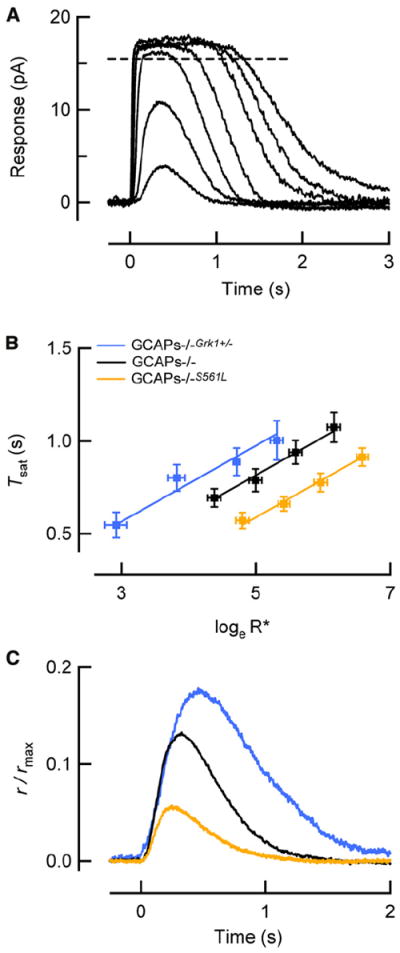
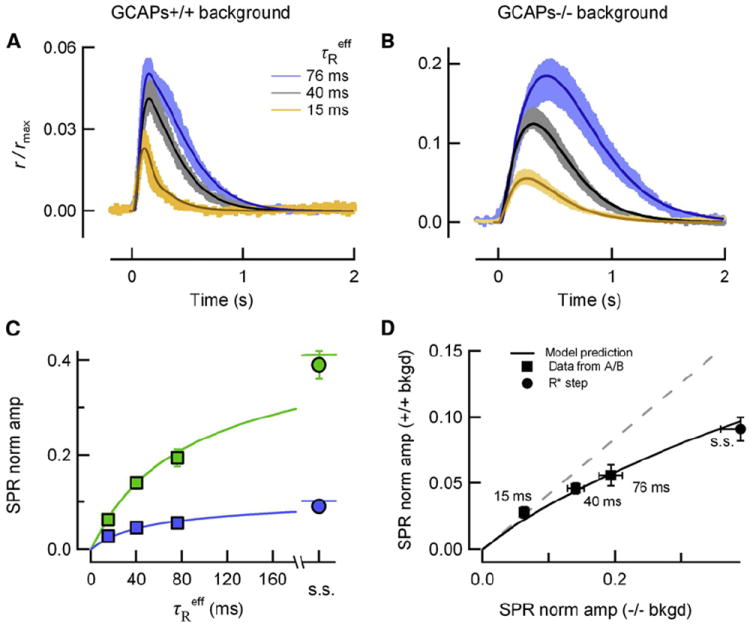
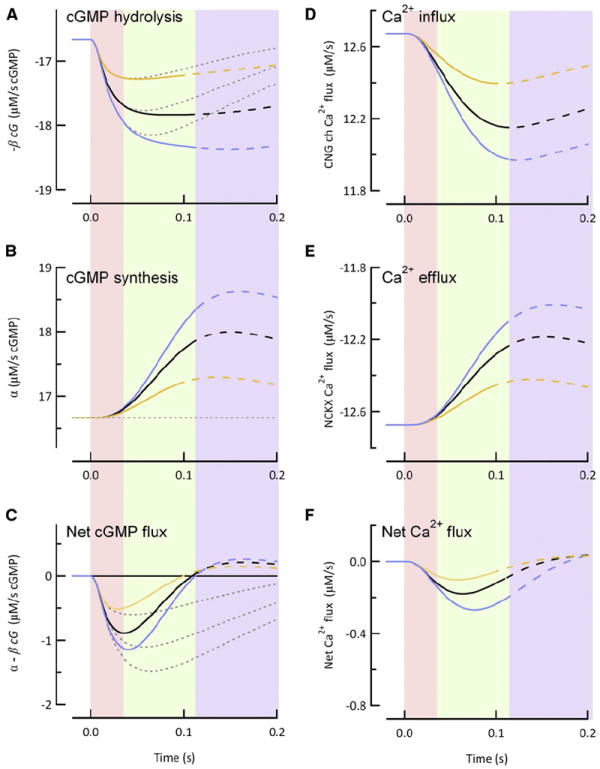
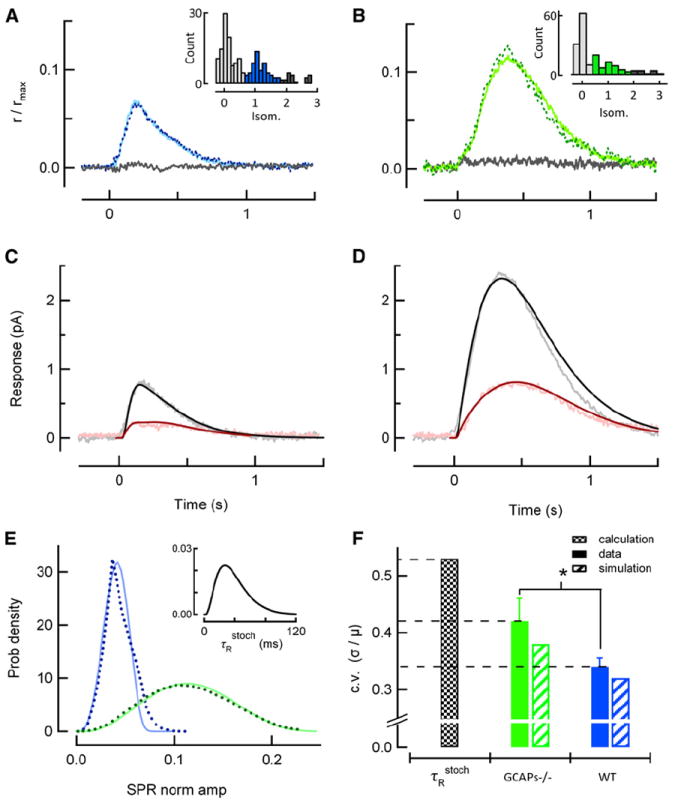
Rod photoreceptors generate amplified, reproducible responses to single photons via a G protein signaling cascade. Surprisingly, genetic perturbations that dramatically alter the deactivation of the principal signal amplifier, the GPCR rhodopsin (R∗), do not much alter the amplitude of single-photon responses (SPRs). These same perturbations, when crossed into a line lacking calcium feedback regulation of cGMP synthesis, produced much larger alterations in SPR amplitudes. Analysis of SPRs from rods with and without feedback reveal that the consequences of trial-to-trial fluctuations in R∗ lifetime in normal rods are also dampened by feedback regulation of cGMP synthesis. Thus, calcium feedback trumps the mechanisms of R∗ deactivation in determining the SPR amplitude, attenuating responses arising from longer R∗ lifetimes to a greater extent than those arising from shorter ones. As a result, rod SPRs achieve a more stereotyped amplitude, a characteristic considered important for reliable transmission through the visual system.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Multiple steps of phosphorylation of activated rhodopsin can account for the reproducibility of vertebrate rod single-photon responses.J Gen Physiol. 2003 Oct;122(4):419-44. doi: 10.1085/jgp.200308832. Epub 2003 Sep 15. J Gen Physiol. 2003. PMID: 12975449 Free PMC article.
-
Constitutive excitation by Gly90Asp rhodopsin rescues rods from degeneration caused by elevated production of cGMP in the dark.J Neurosci. 2007 Aug 15;27(33):8805-15. doi: 10.1523/JNEUROSCI.2751-07.2007. J Neurosci. 2007. PMID: 17699662 Free PMC article.
-
Dynamics of cyclic GMP synthesis in retinal rods.Neuron. 2002 Sep 26;36(1):81-91. doi: 10.1016/s0896-6273(02)00911-x. Neuron. 2002. PMID: 12367508
-
Tuning outer segment Ca2+ homeostasis to phototransduction in rods and cones.Adv Exp Med Biol. 2002;514:179-203. doi: 10.1007/978-1-4615-0121-3_11. Adv Exp Med Biol. 2002. PMID: 12596922 Review.
-
How photons start vision.Proc Natl Acad Sci U S A. 1996 Jan 23;93(2):560-5. doi: 10.1073/pnas.93.2.560. Proc Natl Acad Sci U S A. 1996. PMID: 8570595 Free PMC article. Review.
Cited by
-
Analysis of dim-light responses in rod and cone photoreceptors with altered calcium kinetics.J Math Biol. 2023 Oct 12;87(5):69. doi: 10.1007/s00285-023-02005-4. J Math Biol. 2023. PMID: 37823947 Free PMC article.
-
The evolution of rod photoreceptors.Philos Trans R Soc Lond B Biol Sci. 2017 Apr 5;372(1717):20160074. doi: 10.1098/rstb.2016.0074. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28193819 Free PMC article. Review.
-
Bright flash response recovery of mammalian rods in vivo is rate limited by RGS9.J Gen Physiol. 2017 Apr 3;149(4):443-454. doi: 10.1085/jgp.201611692. Epub 2017 Mar 16. J Gen Physiol. 2017. PMID: 28302678 Free PMC article.
-
Temporal Transformation of the Rod Single-Photon Response in the Retinal Circuitry.Invest Ophthalmol Vis Sci. 2025 Feb 3;66(2):52. doi: 10.1167/iovs.66.2.52. Invest Ophthalmol Vis Sci. 2025. PMID: 39976956 Free PMC article.
-
cGMP in mouse rods: the spatiotemporal dynamics underlying single photon responses.Front Mol Neurosci. 2015 Mar 4;8:6. doi: 10.3389/fnmol.2015.00006. eCollection 2015. Front Mol Neurosci. 2015. PMID: 25788876 Free PMC article. Review.
References
-
- Burns ME, Mendez A, Chen J, Baylor DA. Dynamics of cyclic GMP synthesis in retinal rods. Neuron. 2002;36:81–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
