

Review
doi: 10.1016/j.pt.2012.09.004.
Epub 2012 Oct 16.
Host targeting of virulence determinants and phosphoinositides in blood stage malaria parasites
Affiliations
- PMID: 23084821
- PMCID: PMC3501602
- DOI: 10.1016/j.pt.2012.09.004
Item in Clipboard
Review
Host targeting of virulence determinants and phosphoinositides in blood stage malaria parasites
Trends Parasitol.
2012 Dec.
Abstract
Blood stage malaria parasites target a 'secretome' of hundreds of proteins including virulence determinants containing a host (cell) targeting (HT) signal, to human erythrocytes. Recent studies reveal that the export mechanism is due to the HT signal binding to the lipid phosphatidylinositol-3-phosphate [PI(3)P] in the parasite endoplasmic reticulum (ER). An aspartic protease plasmepsin V which cleaves a specialized form of the HT signal was previously thought to be the export mechanism, but is now recognized as a dedicated peptidase that cleaves the signal anchor subsequent to PI(3)P binding. We discuss a model of PI(3)P-dependent targeting and PI(3)P biology of a major human pathogen.
Published by Elsevier Ltd.
Figures
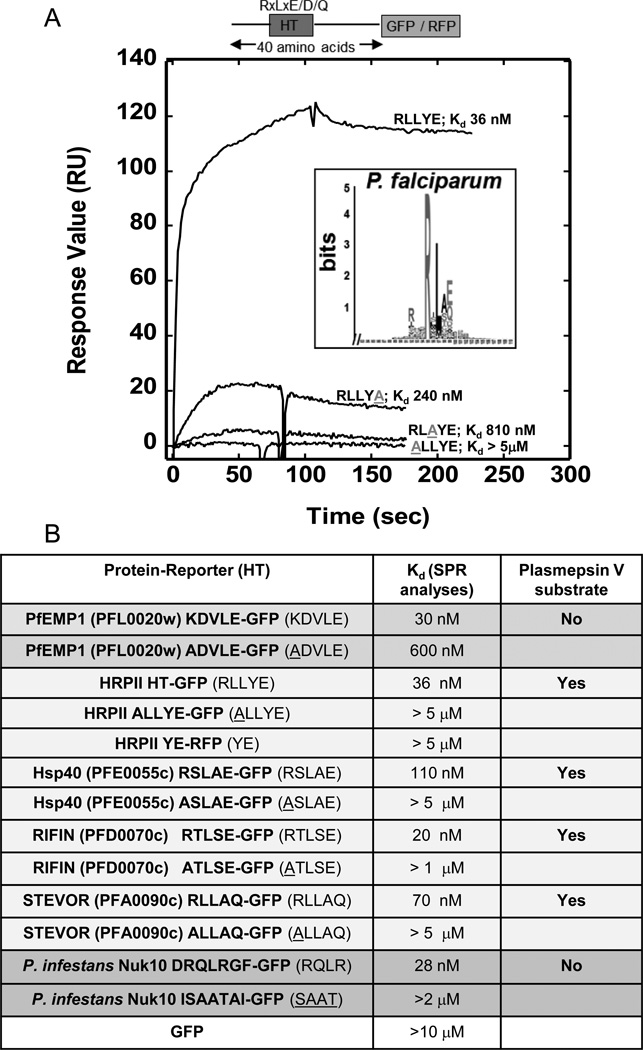
Lipid binding properties of HT signals. (a) Logo and Kd values for each recombinant form of PfHRPII reveals PI(3)P binding with nanomolar affinity and amino acid specificities displayed by HT-mediated export [4]. Sequence logo is derived from HT signal of P. falciparum secretory proteins [10]. Amino acids are represented by one-letter abbreviations. Height of amino acids indicates their frequency at that position. SPR analysis demonstrates quantitative lipid binding of HT/RLLYE-GFP, ALLYE-GFP, RLAYE-GFP, and RLLYA-GFP for PI(3)P containing vesicles. 1 M of each construct was injected over a POPC:POPE:PI(3)P (75:20:5) surface using POPC:POPE (80:20) as a control. The control response was subtracted from each active surface to yield the displayed sensorgrams. Adapted from [4]. (b) A summary of PI(3)P binding properties and plasmepsin V susceptibility of HT motifs of P. falciparum PfEMP1, PfHRPII, PfHsp40, RIFIN, STEVOR as well as P. infestans Nuk10. Adapted from [4], [20], [21].
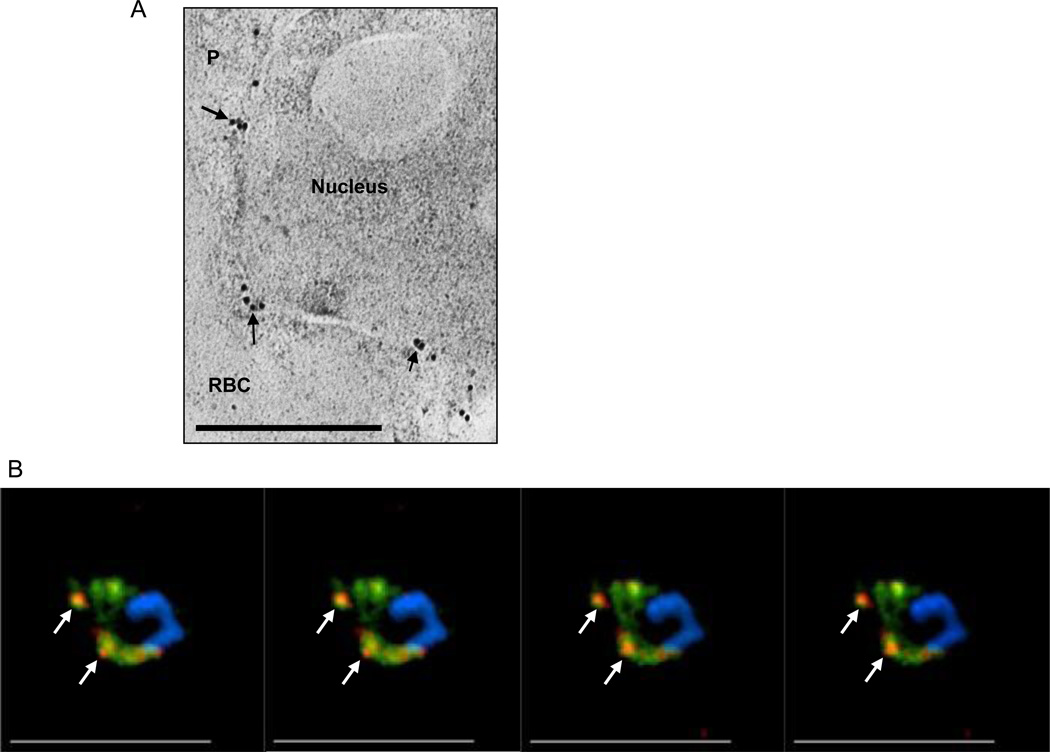
PI(3)P is detected in P. falciparum endoplasmic reticulum. (a) Immuno-electronmicroscopy showing localization of secretory EEA1WT-mCherry in reticular perinuclear membranes characteristic of the ER. Thin sections, probed with anti-mCherry and secondary antibody gold conjugates (10 nm) show label concentrated in membrane regions emerging from reticular membrane apposed to the nucleus (arrows), consistent with localization in the ER. Scale bar, 0.5 µm. [4]. (b) Successive single optical z-sections of a precursor ER form of a secretome protein PfHRPII (pPfHRPII; green) prior to cleavage by plasmepsin V and its colocalization with PI(3)P regions (red). Subsets of prominent areas of colocalization are indicated by arrows. Abbreviations: RBC, red blood cell; P, parasite.
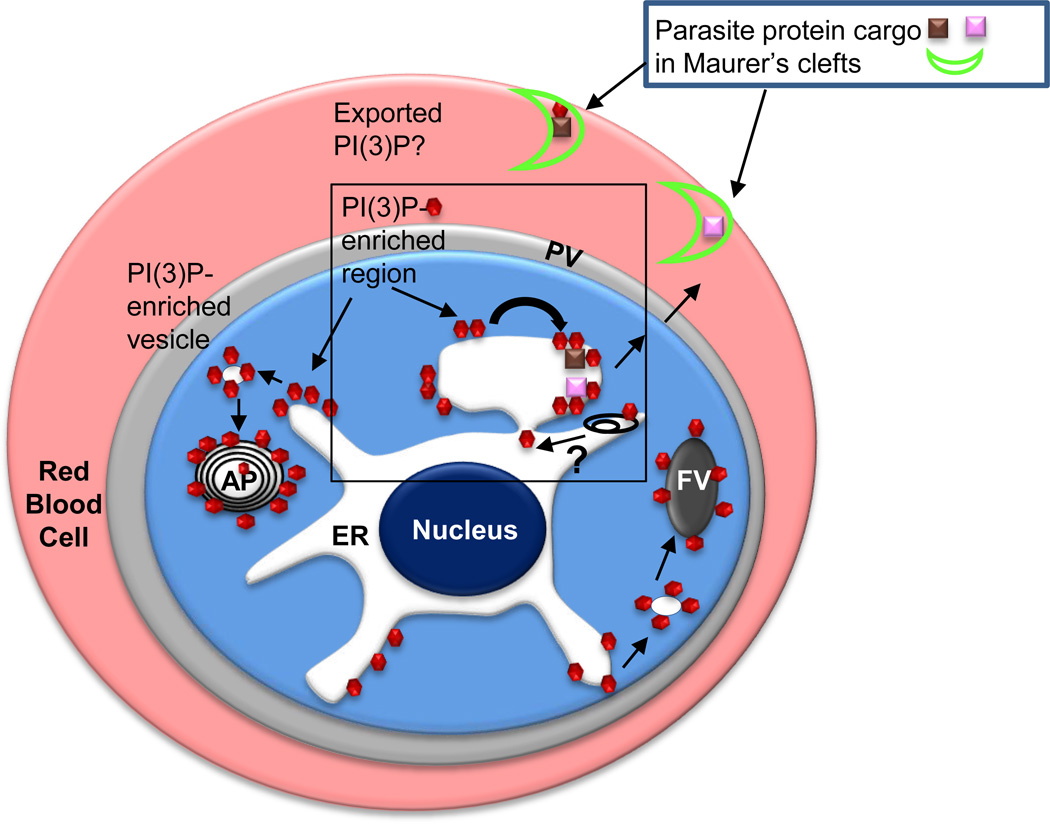
Secretory and cytoplasmic distribution of PI(3)P in P. falciparum-infected erythrocytes. Since P. falciparum encodes for a single PI3 kinase, PI(3)P (red hexagons) is expected to be synthesized on the cytoplasmic face of membranes. PI(3)P is reported to be prominently associated with the cytoplasmic face of the apicoplast and the food vacuole. PI(3)P is reported to be essential for hemoglobin uptake from the red cell into the parasite food vacuole (FV) as well as apicoplast (AP) biogenesis [25]. As highlighted in Box 1, PI(3)P is also detected in the lumen of the ER, suggesting it undergoes trans bilayer ‘flip-flop’ from the cytoplasmic face (black curved arrow) either through the action of a ‘flippase’ or possibly internalization of PI(3)P bound by the HT motif as the nascent peptide translocates across the ER membrane. Autophagy (represented by black endomembranes) may also provide a route of uptake of cytoplasmic PI(3)P but whether this releases PI(3)P with correct topology in the ER lumen is unknown (depicted by a question mark). Proteins (pink and brown squares) bound to PI(3)P through an HT motif are exported Maurer’s clefts (bright green crescent) in the host erythrocyte. If the HT signal is not cleaved, PI(3)P may be exported to the host erythrocyte, but for the vast majority of secretome proteins, PI(3)P is not exported to the host erythrocyte.
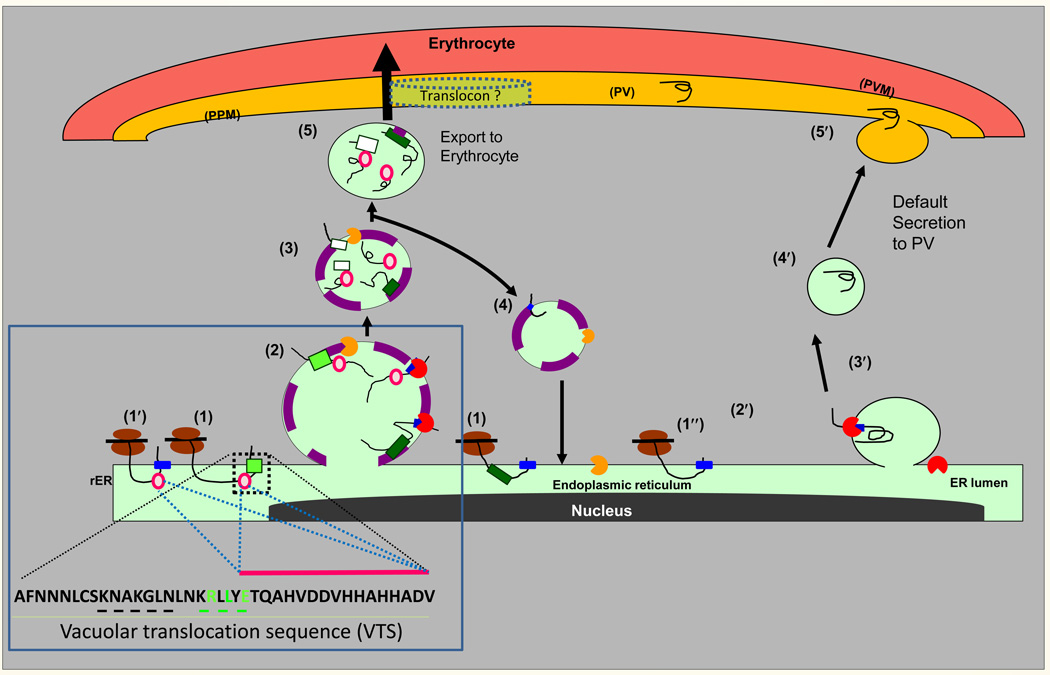
A model for PI(3)P-dependent and- independent export from the ER of P. falciparum-infected erythrocytes. Proteins containing the malarial HT-signal (light green square) or the oomycete HT signal (dark green square) are co-translationally inserted via their signal anchor sequences (blue squares) into the ER membrane (step 1). The HT signals recognize the lipid PI(3)P (purple) enriched in regions of the ER (step 2) and may occur co-translationally (not shown). Secretory proteins with a plasmepsin V-refractory HT signal, are cleaved by signal peptidase (red pac-man) but remain associated to PI(3)P. Proteins with the malarial HT signal are cleaved by plasmepsin V (orange pac-man, step 3) which also destroys the PI(3)P binding signal and thus is likely to occur in a newly pinched off vesicle or one whose contents do not freely diffuse with the ER. In addition, highly charged peptide sequences (pink circles) downstream of the HT signal capable of PI(3)P-independent export may also facilitate association with the host targeted pathway despite absence/cleavage of the HT motif (1’ and steps 2 and later). Plasmepsin V and PI(3)P are recycled back to the ER (step 4), while cargo targeted to the erythrocyte moves forward across the parasite plasma membrane (PPM), and PVM (step 5). In default secretion, secretory proteins are co-translationally translocated into the ER (step 1′), the signal sequence (blue square) is cleaved by signal peptidase (red pac-man; steps 2′ and 3′) and protein is delivered through vesicular intermediates to PPM, and released into the PV (steps 4′ and 5′). Steps 1–5 have ~400 predicted cargo proteins exported to the erythrocyte and thus likely the dominant pathway of protein export to the erythrocyte. The role of the Golgi in these pathways is not known. A translocon has been proposed in export to the erythrocyte, but how it recognizes HT signals lost in the ER is unknown. Adapted from [4]. Box 1 describes the vacuolar translocation sequence of PfHRPII and a summary of regions/residues needed for export as well as PI(3)P binding. Shown in green are conserved HT motif residues R, L and E known to contribute to both PI(3)P binding and protein export to the erythrocyte. Substitution of sequences KNAKGLN upstream of the HT motif (underlined in black), also blocked export [30], but have not been tested for PI(3)P binding. Pink circles and pink bar indicating charged amino acids downstream of the HT motif enable export independent of PI(3)P, however, the E residue marked in green contributes to PI(3)P binding in the intact HT motif (see Figure 1). Substitution of HHAHHA sequence in repeat sequences does not block export [30].
Similar articles
-
Endoplasmic reticulum PI(3)P lipid binding targets malaria proteins to the host cell.Cell. 2012 Jan 20;148(1-2):201-12. doi: 10.1016/j.cell.2011.10.051. Cell. 2012. PMID: 22265412 Free PMC article.
-
PI(3)P-independent and -dependent pathways function together in a vacuolar translocation sequence to target malarial proteins to the host erythrocyte.Mol Biochem Parasitol. 2012 Oct;185(2):106-13. doi: 10.1016/j.molbiopara.2012.07.004. Epub 2012 Jul 22. Mol Biochem Parasitol. 2012. PMID: 22828070 Free PMC article.
-
Plasmepsin V licenses Plasmodium proteins for export into the host erythrocyte.Nature. 2010 Feb 4;463(7281):632-6. doi: 10.1038/nature08726. Nature. 2010. PMID: 20130644 Free PMC article.
-
Eukaryotic virulence determinants utilize phosphoinositides at the ER and host cell surface.Trends Microbiol. 2013 Mar;21(3):145-56. doi: 10.1016/j.tim.2012.12.004. Epub 2013 Jan 30. Trends Microbiol. 2013. PMID: 23375057 Free PMC article. Review.
-
Cerebral malaria pathogenesis: revisiting parasite and host contributions.Future Microbiol. 2012 Feb;7(2):291-302. doi: 10.2217/fmb.11.155. Future Microbiol. 2012. PMID: 22324996 Review.
Cited by
-
Genome analysis of Excretory/Secretory proteins in Taenia solium reveals their Abundance of Antigenic Regions (AAR).Sci Rep. 2015 May 19;5:9683. doi: 10.1038/srep09683. Sci Rep. 2015. PMID: 25989346 Free PMC article.
-
Why do malaria parasites increase host erythrocyte permeability?Trends Parasitol. 2014 Mar;30(3):151-9. doi: 10.1016/j.pt.2014.01.003. Epub 2014 Feb 5. Trends Parasitol. 2014. PMID: 24507014 Free PMC article. Review.
-
Whole genome sequencing identifies novel mutations in malaria parasites resistant to artesunate (ATN) and to ATN + mefloquine combination.Front Cell Infect Microbiol. 2024 Mar 1;14:1353057. doi: 10.3389/fcimb.2024.1353057. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38495651 Free PMC article.
-
The Plasmodium berghei translocon of exported proteins reveals spatiotemporal dynamics of tubular extensions.Sci Rep. 2015 Jul 29;5:12532. doi: 10.1038/srep12532. Sci Rep. 2015. PMID: 26219962 Free PMC article.
-
The systematic analysis of protein-lipid interactions comes of age.Nat Rev Mol Cell Biol. 2015 Dec;16(12):753-61. doi: 10.1038/nrm4080. Epub 2015 Oct 28. Nat Rev Mol Cell Biol. 2015. PMID: 26507169 Review.
References
-
- Collins WE. Plasmodium knowlesi: a malaria parasite of monkeys and humans. Annual review of entomology. 2012;57:107–121. - PubMed
-
- Miller LH, et al. The pathogenic basis of malaria. Nature. 2002;415:673–679. - PubMed
-
- Haldar K, Mohandas N. Erythrocyte remodeling by malaria parasites. Curr Opin Hematol. 2007;14:203–209. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
