

Innate lymphoid cells: balancing immunity, inflammation, and tissue repair in the intestine
- PMID: 23084914
- PMCID: PMC3584448
- DOI: 10.1016/j.chom.2012.10.003
Innate lymphoid cells: balancing immunity, inflammation, and tissue repair in the intestine
Abstract
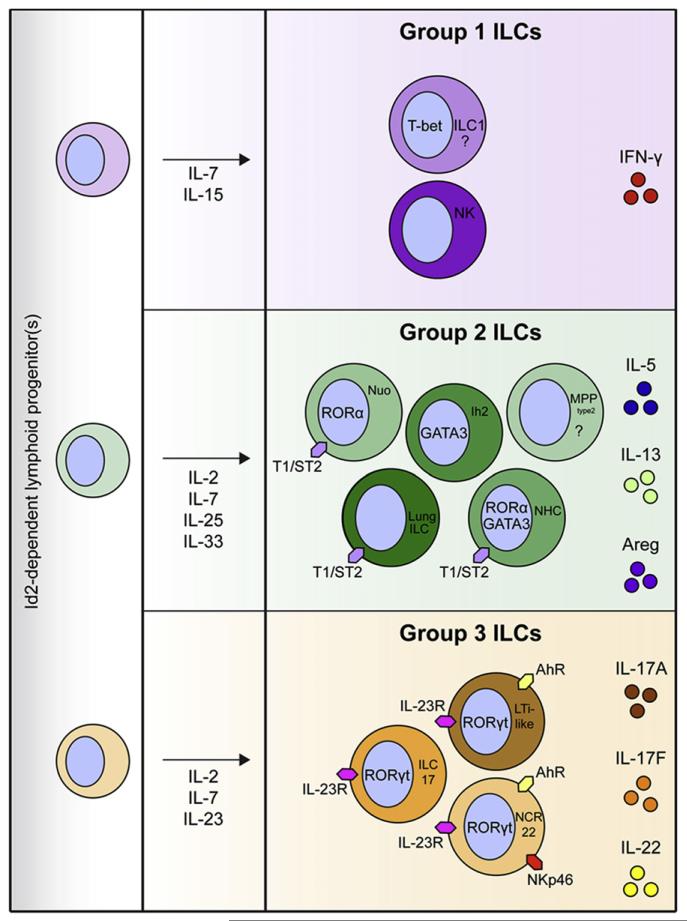
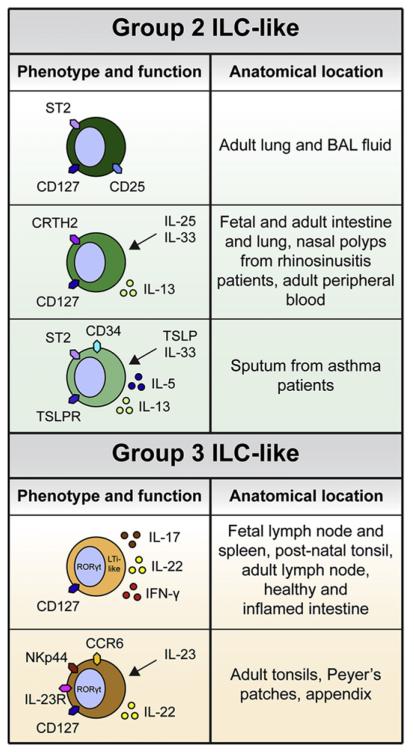
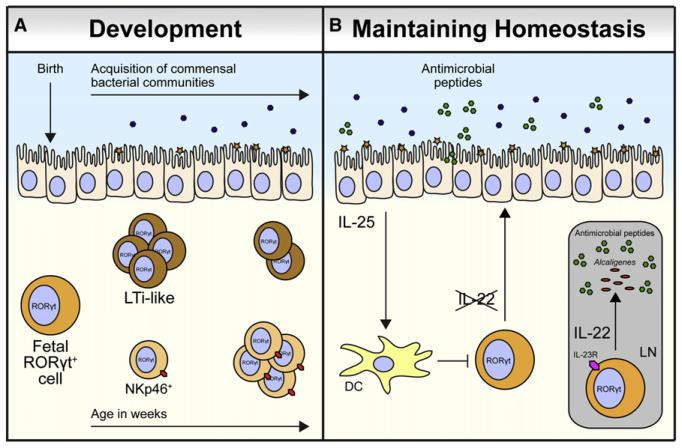
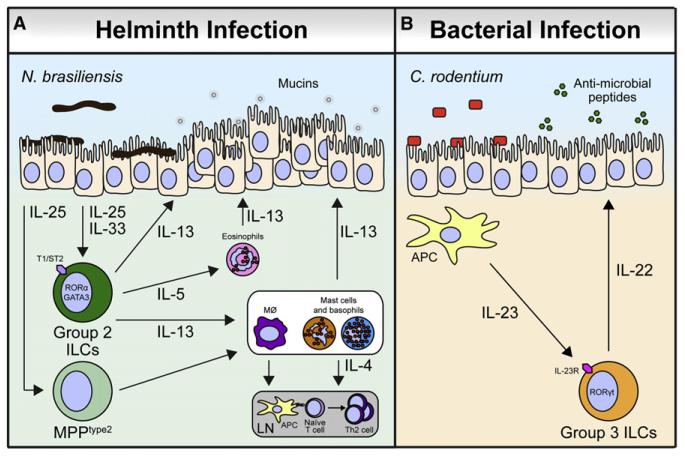
Innate lymphoid cells (ILCs) are a recently described group of innate immune cells that can regulate immunity, inflammation, and tissue repair in multiple anatomical compartments, particularly the barrier surfaces of the skin, airways, and intestine. Broad categories of ILCs have been defined based on transcription factor expression and the ability to produce distinct patterns of effector molecules. Recent studies have revealed that ILC populations can regulate commensal bacterial communities, contribute to resistance to helminth and bacterial pathogens, promote inflammation, and orchestrate tissue repair and wound healing. This review will examine the phenotype and function of murine and human ILCs and discuss the critical roles these innate immune cells play in health and disease.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

References
-
- Ahern PP, Izcue A, Maloy KJ, Powrie F. The interleukin-23 axis in intestinal inflammation. Immunol. Rev. 2008;226:147–159. - PubMed
-
- Allakhverdi Z, Comeau MR, Smith DE, Toy D, Endam LM, Desrosiers M, Liu YJ, Howie KJ, Denburg JA, Gauvreau GM, Delespesse G. CD34+ hemopoietic progenitor cells are potent effectors of allergic inflammation. J. Allergy Clin. Immunol. 2009;123:472–478. - PubMed
-
- Barlow JL, Bellosi A, Hardman CS, Drynan LF, Wong SH, Cruickshank JP, McKenzie AN. Innate IL-13-producing nuocytes arise during allergic lung inflammation and contribute to airways hyperreactivity. J. Allergy Clin. Immunol. 2012;129:191–198. e1–e4. - PubMed
Publication types
MeSH terms
Grants and funding
- R01 AI074878/AI/NIAID NIH HHS/United States
- T32-AI060516/AI/NIAID NIH HHS/United States
- R01 AI095466/AI/NIAID NIH HHS/United States
- AI061570/AI/NIAID NIH HHS/United States
- F32-AI098365/AI/NIAID NIH HHS/United States
- AI074878/AI/NIAID NIH HHS/United States
- AI087990/AI/NIAID NIH HHS/United States
- T32 AI060516/AI/NIAID NIH HHS/United States
- R21 AI087990/AI/NIAID NIH HHS/United States
- AI095608/AI/NIAID NIH HHS/United States
- R21 AI083480/AI/NIAID NIH HHS/United States
- F32 AI098365/AI/NIAID NIH HHS/United States
- U01 AI095608/AI/NIAID NIH HHS/United States
- R01 AI061570/AI/NIAID NIH HHS/United States
- AI097333/AI/NIAID NIH HHS/United States
- AI083480/AI/NIAID NIH HHS/United States
- R01 AI102942/AI/NIAID NIH HHS/United States
- R01 AI097333/AI/NIAID NIH HHS/United States
- AI095466/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
