

Lipid droplets control the maternal histone supply of Drosophila embryos
- PMID: 23084995
- PMCID: PMC3513403
- DOI: 10.1016/j.cub.2012.09.018
Lipid droplets control the maternal histone supply of Drosophila embryos
Abstract
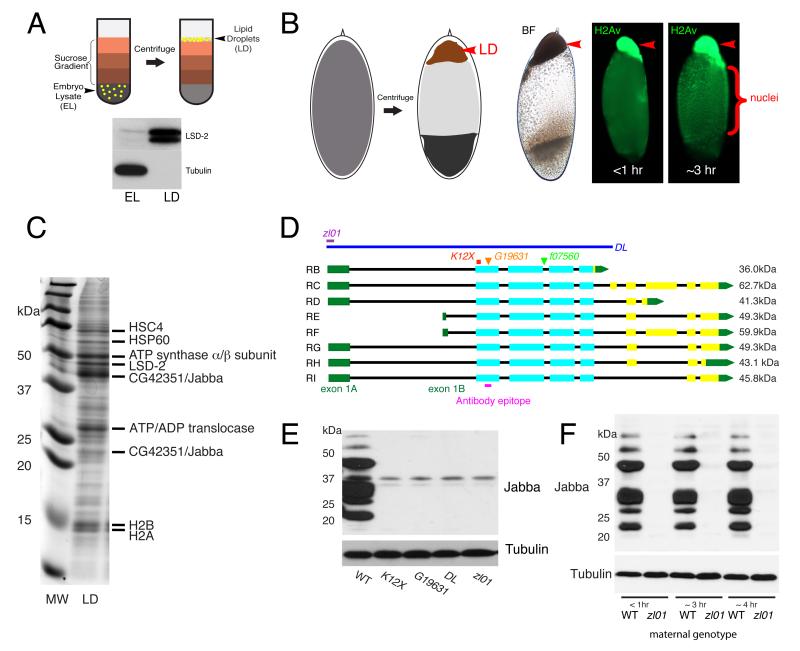
Background: Histones are essential for chromatin packing, yet free histones not incorporated into chromatin are toxic. While in most cells multiple regulatory mechanisms prevent accumulation of excess histones, early Drosophila embryos contain massive extranuclear histone stores, thought to be essential for development. Excess histones H2A, H2B, and H2Av are bound to lipid droplets, ubiquitous fat storage organelles especially abundant in embryos. It has been proposed that sequestration on lipid droplets allows safe transient storage of supernumerary histones.
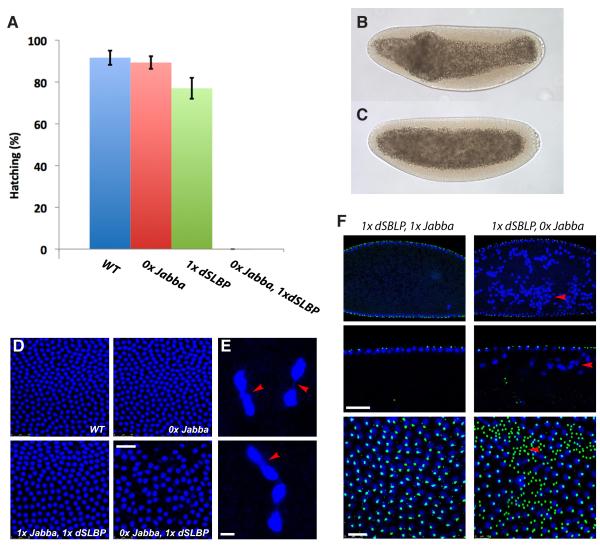
Results: Here, we critically test this sequestration hypothesis. We find that histones are anchored to lipid droplets via the previously uncharacterized protein Jabba: Jabba localizes to droplets, coimmunoprecipitates with histones, and is necessary to recruit histones to droplets. Jabba mutants lack the maternal H2A, H2B, and H2Av deposits altogether; presumably, these deposits are eliminated unless sequestered on droplets. Jabba mutant embryos compensate for this histone deficit by translating maternal histone mRNAs. However, when histone expression is mildly compromised, the maternal histone protein deposits are essential for proper early mitoses and for viability.
Conclusions: A growing number of proteins from other cellular compartments have been found to transiently associate with lipid droplets. Our studies provide the first insight into mechanism and functional relevance of this sequestration. We conclude that sequestration on lipid droplets allows embryos to build up extranuclear histone stores and provides histones for chromatin assembly during times of high demand. This work reveals a novel aspect of histone metabolism and establishes lipid droplets as functional storage sites for unstable or detrimental proteins.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures





Comment in
-
Organelle dynamics: a histone anchor on lipid droplets.Nat Rev Mol Cell Biol. 2012 Dec;13(12):753. doi: 10.1038/nrm3480. Nat Rev Mol Cell Biol. 2012. PMID: 23175278 No abstract available.
References
-
- Gunjan A, Verreault A. A Rad53 kinase-dependent surveillance mechanism that regulates histone protein levels in S. cerevisiae. Cell. 2003;115:537–549. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
