

Replication-dependent irreversible topoisomerase 1 poisoning is responsible for FdUMP[10] anti-leukemic activity
- PMID: 23085462
- PMCID: PMC3660094
- DOI: 10.1016/j.exphem.2012.10.007
Replication-dependent irreversible topoisomerase 1 poisoning is responsible for FdUMP[10] anti-leukemic activity
Abstract
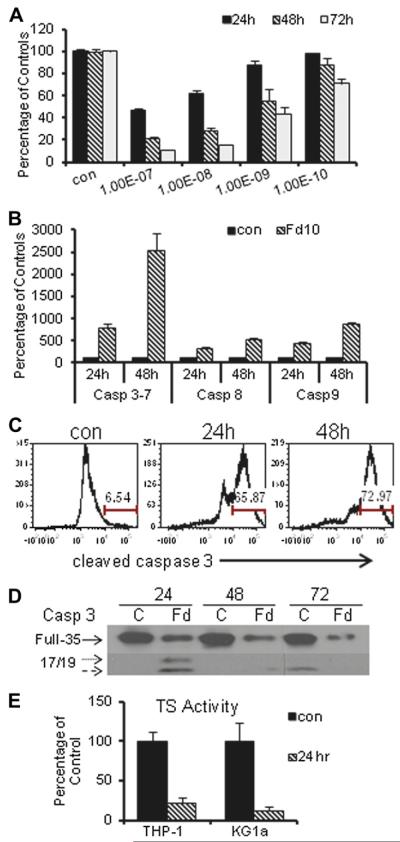
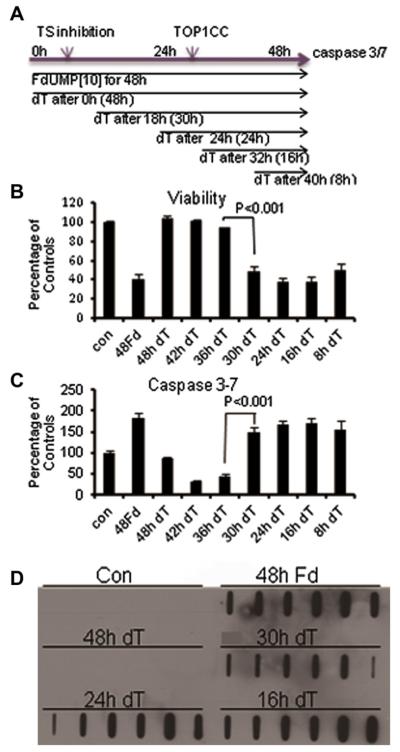
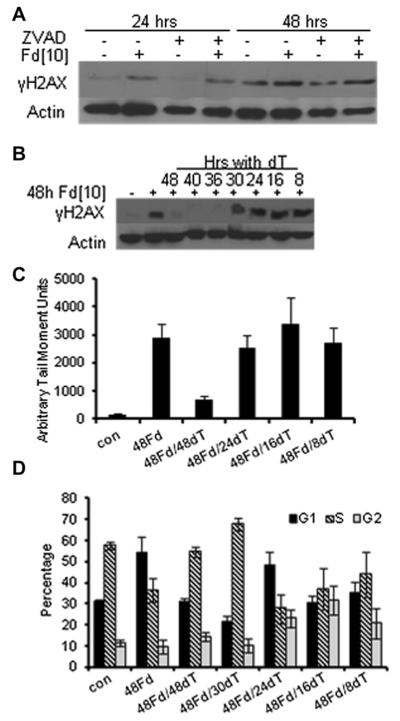
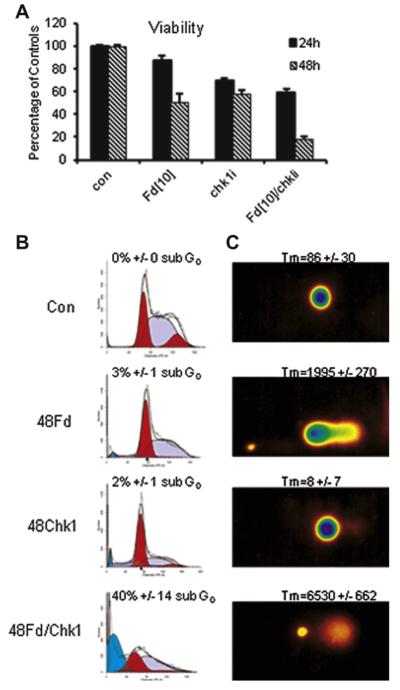
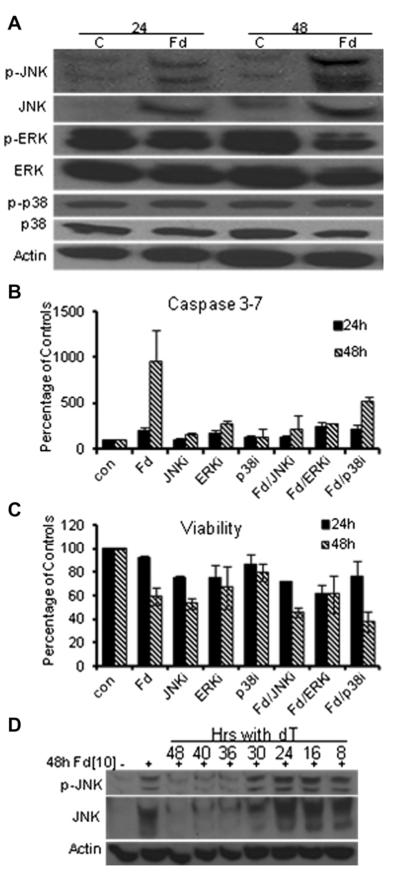
Previous studies have indicated that 5-Fluoro-2'-deoxyuridine-5'-O-monophosphate 10mer (FdUMP[10]) displays strong antileukemic activity through the dual targeting of thymidylate synthase (TS) and DNA topoisomerase 1 (Top1). The present studies were undertaken to clarify the relationship between the induction of a thymineless state and the formation of Top1 cleavage complexes (Top1CC) for inducing cell death and to clarify the role of DNA replication for induction of lethal DNA double-strand breaks (DSBs) in FdUMP[10]-treated acute myeloid leukemia (AML) cells. Human promyelocytic (HL60) and AML (KG1a, Molm13, THP-1) cells were synchronized by serum starvation and treated with FdUMP[10] with thymidine (Thy) rescue. Cells were assayed for TS inhibition, DNA DSBs, Top1CC, and apoptosis to clarify the interrelationship of TS inhibition and Top1CC for cell death. FdUMP[10] induced a thymineless state in AML cells and exogenous Thy administered within the first 18 hours of treatment rescued FdUMP[10]-induced Top1CC formation, γH2AX phosphorylation, and apoptosis induction. Exogenous Thy was not effective after cells had committed to mitosis and undergone cell division in the presence of FdUMP[10]. FdUMP[10] treatment resulted in Chk1 activation, and Chk1 inhibition enhanced FdUMP[10]-induced DNA damage and apoptosis. Jnk-signaling was required for FdUMP[10]-induced apoptosis in promyelocytic HL60 cells and in THP1 cells, but was antiapoptotic in Molm13 and to a lesser extent KG1a AML cells. The results are consistent with FdUMP[10] inducing a thymineless state, leading to misincorporation of FdU into genomic DNA of proliferating cells. Top1CC form in cells upon re-entry into S-phase, resulting in DNA double-strand breaks, and initiating apoptotic signaling that can be either muted or enhanced by Jnk-signaling depending on cell type.
Copyright © 2013 ISEH - Society for Hematology and Stem Cells. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Roboz GJ. Novel approaches to the treatment of acute myeloid leukemia. Hematology Am Soc Hematol Educ Program. 2011;2011:43–50. - PubMed
-
- Buchner T, Berdel WE, Wormann B, et al. Treatment of older patients with AML. Crit Rev Oncol Hematol. 2005;56:247–259. - PubMed
-
- Robak T, Wierzbowska A. Current and emerging therapies for acute myeloid leukemia. Clin Ther. 2009;31:2349–2370. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
