

Temporal and spatial adaptation of transient responses to local features
- PMID: 23087617
- PMCID: PMC3474938
- DOI: 10.3389/fncir.2012.00074
Temporal and spatial adaptation of transient responses to local features
Abstract
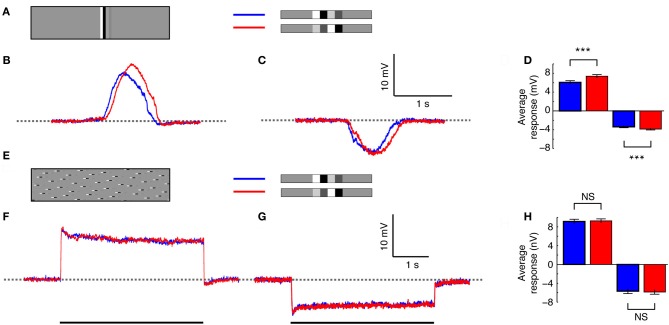
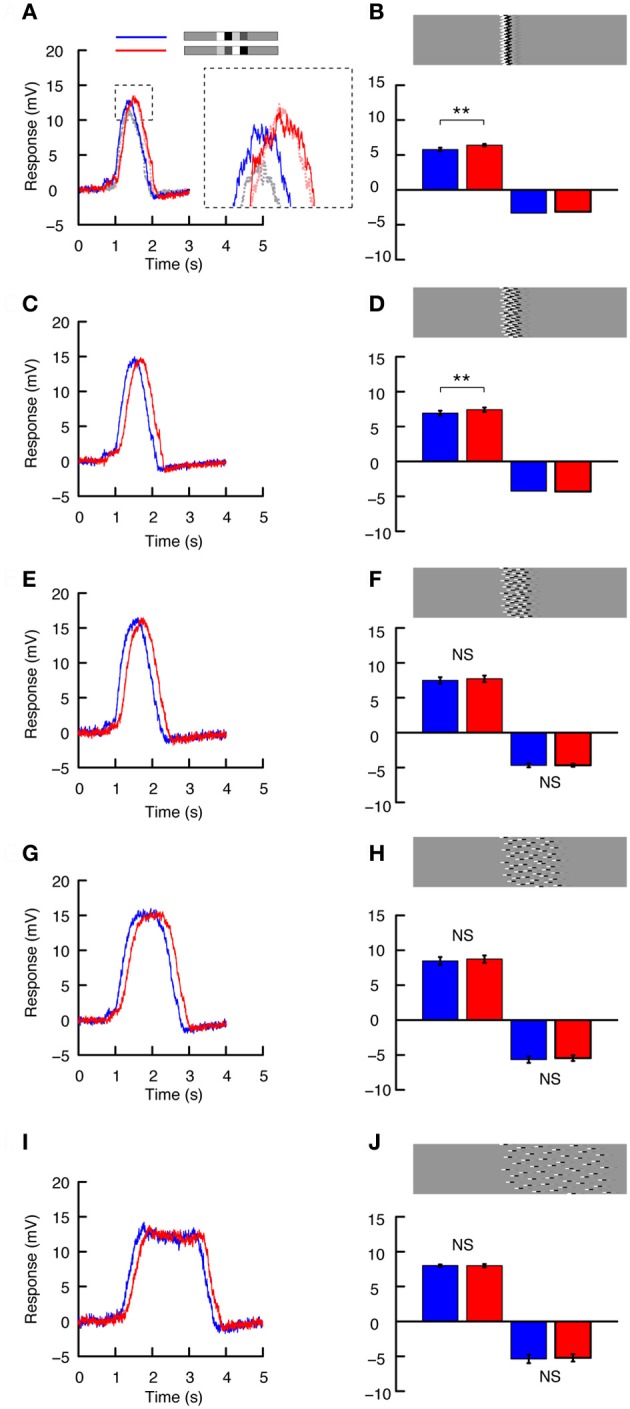
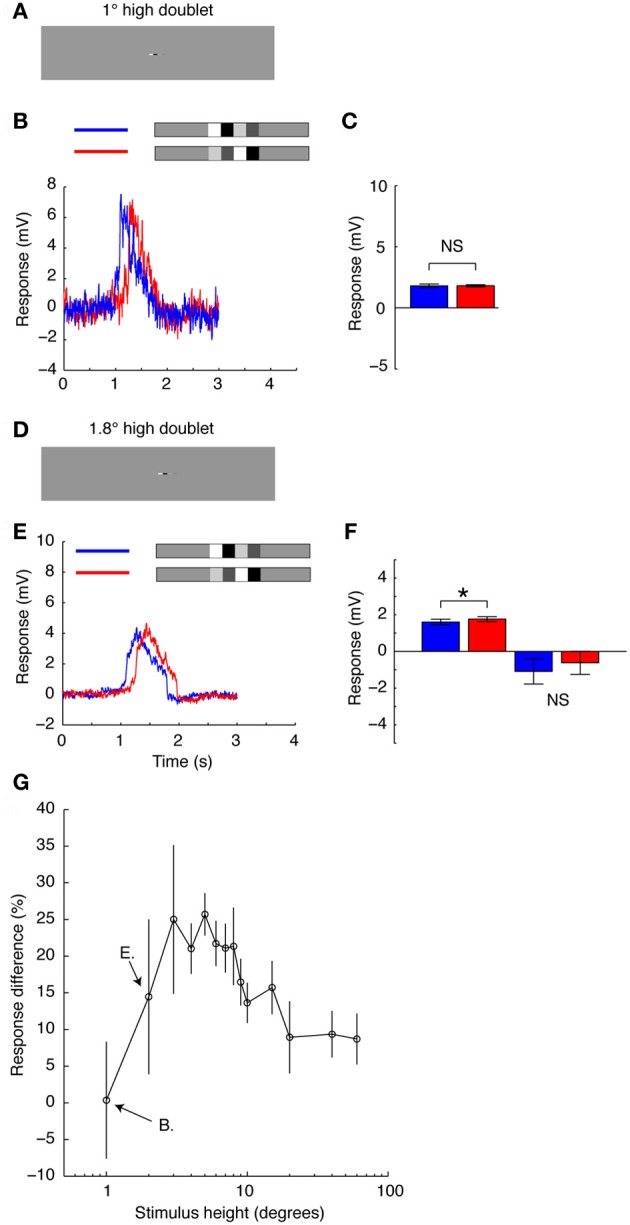
Interpreting visual motion within the natural environment is a challenging task, particularly considering that natural scenes vary enormously in brightness, contrast and spatial structure. The performance of current models for the detection of self-generated optic flow depends critically on these very parameters, but despite this, animals manage to successfully navigate within a broad range of scenes. Within global scenes local areas with more salient features are common. Recent work has highlighted the influence that local, salient features have on the encoding of optic flow, but it has been difficult to quantify how local transient responses affect responses to subsequent features and thus contribute to the global neural response. To investigate this in more detail we used experimenter-designed stimuli and recorded intracellularly from motion-sensitive neurons. We limited the stimulus to a small vertically elongated strip, to investigate local and global neural responses to pairs of local "doublet" features that were designed to interact with each other in the temporal and spatial domain. We show that the passage of a high-contrast doublet feature produces a complex transient response from local motion detectors consistent with predictions of a simple computational model. In the neuron, the passage of a high-contrast feature induces a local reduction in responses to subsequent low-contrast features. However, this neural contrast gain reduction appears to be recruited only when features stretch vertically (i.e., orthogonal to the direction of motion) across at least several aligned neighboring ommatidia. Horizontal displacement of the components of elongated features abolishes the local adaptation effect. It is thus likely that features in natural scenes with vertically aligned edges, such as tree trunks, recruit the greatest amount of response suppression. This property could emphasize the local responses to such features vs. those in nearby texture within the scene.
Keywords: EMD; insect vision; local contrast sensitivity; motion adaptation; motion detection; salient feature; spatial integration.
Figures
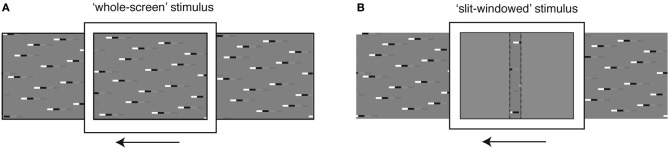
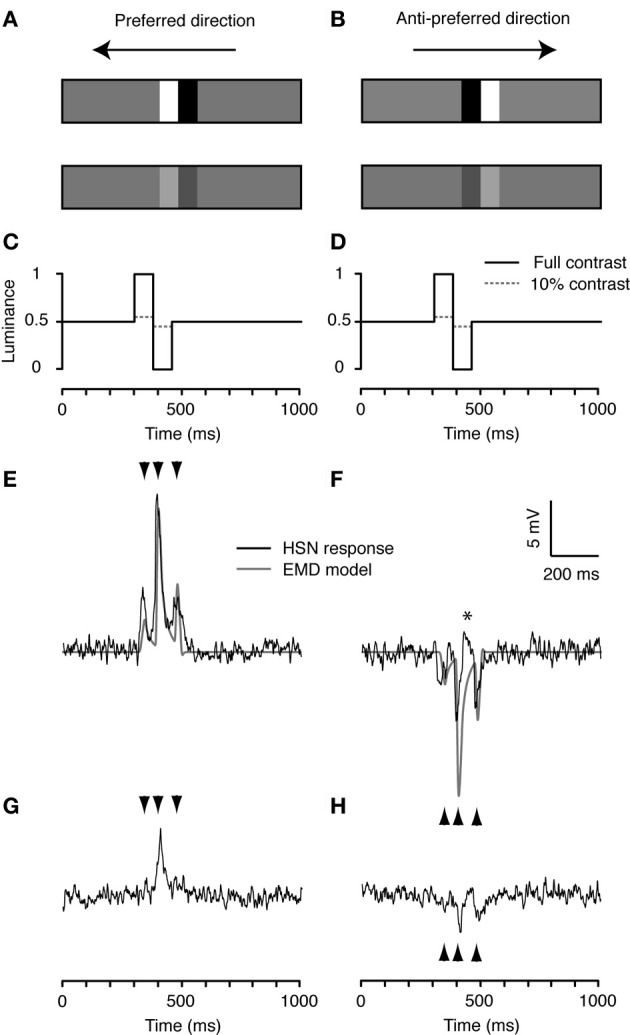
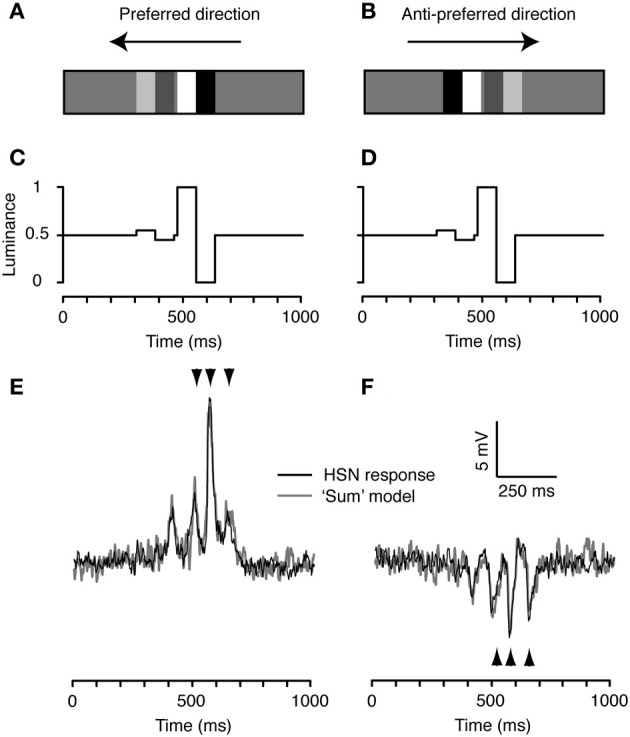
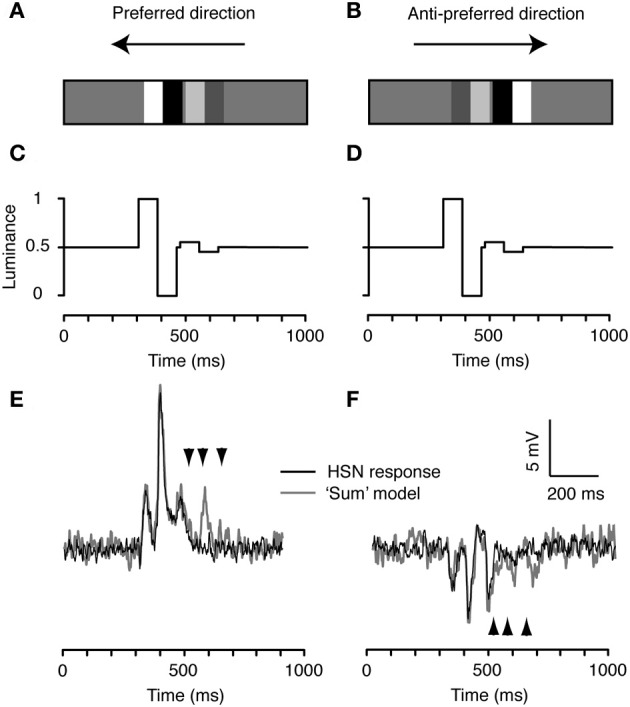
Similar articles
-
Local and global responses of insect motion detectors to the spatial structure of natural scenes.J Vis. 2011 Dec 27;11(14):20. doi: 10.1167/11.14.20. J Vis. 2011. PMID: 22201615
-
Disentangling of Local and Wide-Field Motion Adaptation.Front Neural Circuits. 2021 Aug 31;15:713285. doi: 10.3389/fncir.2021.713285. eCollection 2021. Front Neural Circuits. 2021. PMID: 34531728 Free PMC article.
-
Influence of environmental information in natural scenes and the effects of motion adaptation on a fly motion-sensitive neuron during simulated flight.Biol Open. 2014 Dec 12;4(1):13-21. doi: 10.1242/bio.20149449. Biol Open. 2014. PMID: 25505148 Free PMC article.
-
Motion adaptation and the velocity coding of natural scenes.Curr Biol. 2010 Jun 8;20(11):994-9. doi: 10.1016/j.cub.2010.03.072. Epub 2010 May 27. Curr Biol. 2010. PMID: 20537540
-
The spatial resolutions of the apposition compound eye and its neuro-sensory feature detectors: observation versus theory.J Insect Physiol. 2005 Mar;51(3):243-66. doi: 10.1016/j.jinsphys.2004.11.018. J Insect Physiol. 2005. PMID: 15749108 Review.
Cited by
-
Image statistics of the environment surrounding freely behaving hoverflies.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2019 Jun;205(3):373-385. doi: 10.1007/s00359-019-01329-1. Epub 2019 Apr 1. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2019. PMID: 30937518 Free PMC article.
-
Spatial Encoding of Translational Optic Flow in Planar Scenes by Elementary Motion Detector Arrays.Sci Rep. 2018 Apr 11;8(1):5821. doi: 10.1038/s41598-018-24162-z. Sci Rep. 2018. PMID: 29643402 Free PMC article.
-
Enhancement of prominent texture cues in fly optic flow processing.Front Neural Circuits. 2012 Oct 29;6:78. doi: 10.3389/fncir.2012.00078. eCollection 2012. Front Neural Circuits. 2012. PMID: 23112763 Free PMC article. No abstract available.
-
Spatio-temporal dynamics of impulse responses to figure motion in optic flow neurons.PLoS One. 2015 May 8;10(5):e0126265. doi: 10.1371/journal.pone.0126265. eCollection 2015. PLoS One. 2015. PMID: 25955416 Free PMC article.
-
Localized and Long-Lasting Adaptation in Dragonfly Target-Detecting Neurons.eNeuro. 2024 Sep 20;11(9):ENEURO.0036-24.2024. doi: 10.1523/ENEURO.0036-24.2024. Print 2024 Sep. eNeuro. 2024. PMID: 39256041 Free PMC article.
References
-
- Arnett D. W. (1972). Spatial and temporal integration properties of units in first optic ganglion of dipterans. J. Neurophysiol. 35, 429–444 - PubMed
LinkOut - more resources
Full Text Sources