

Systematic evaluation of the dependence of deoxyribozyme catalysis on random region length
- PMID: 23088677
- PMCID: PMC3518697
- DOI: 10.1021/co300111f
Systematic evaluation of the dependence of deoxyribozyme catalysis on random region length
Abstract
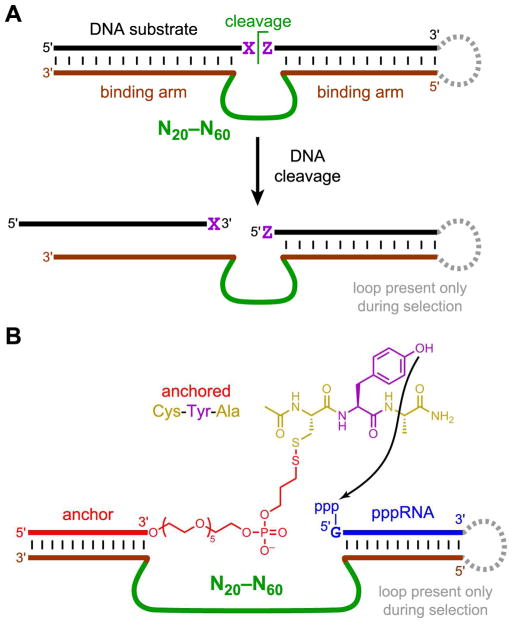
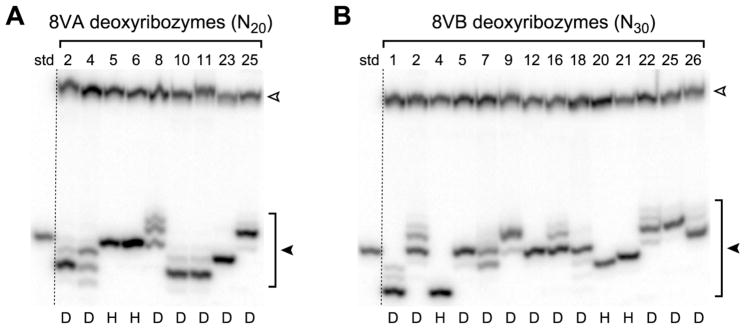
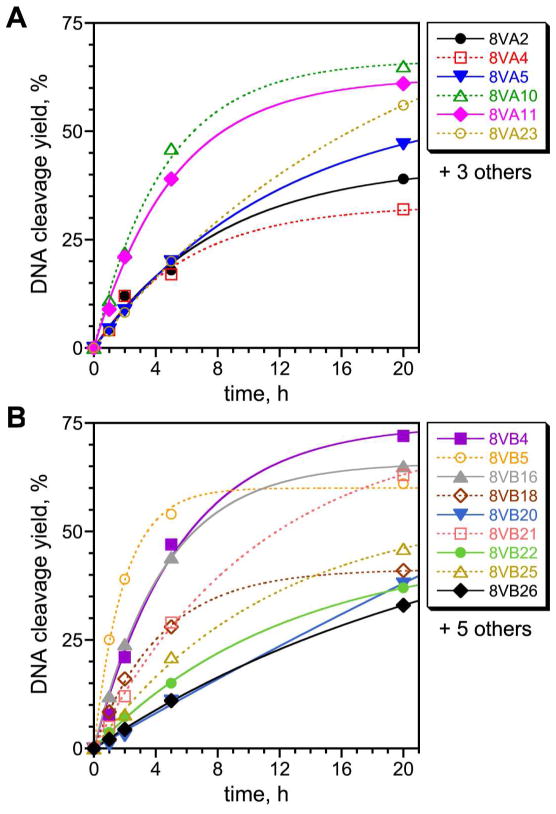
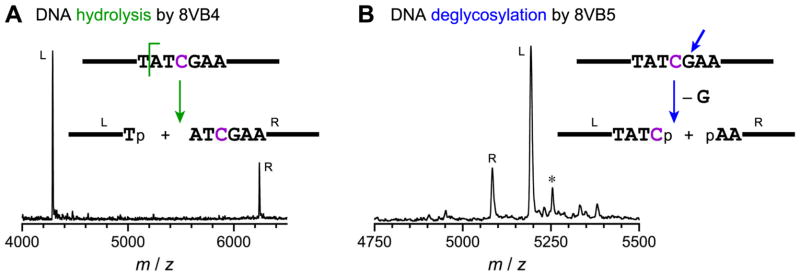
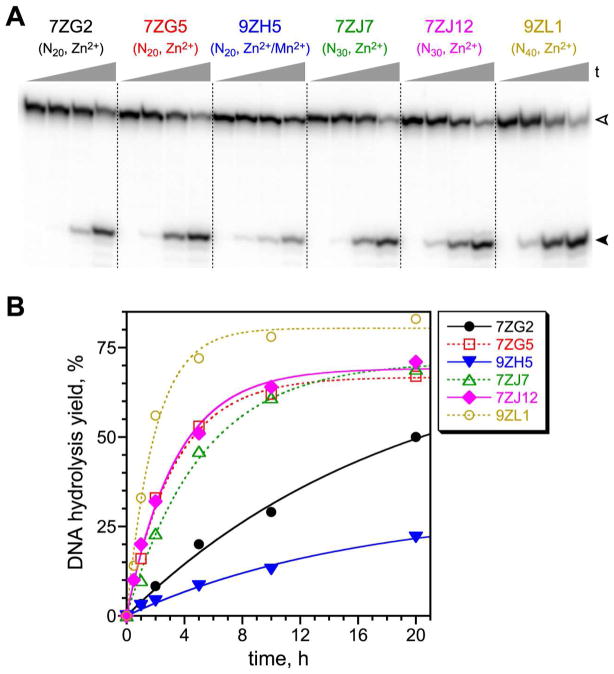
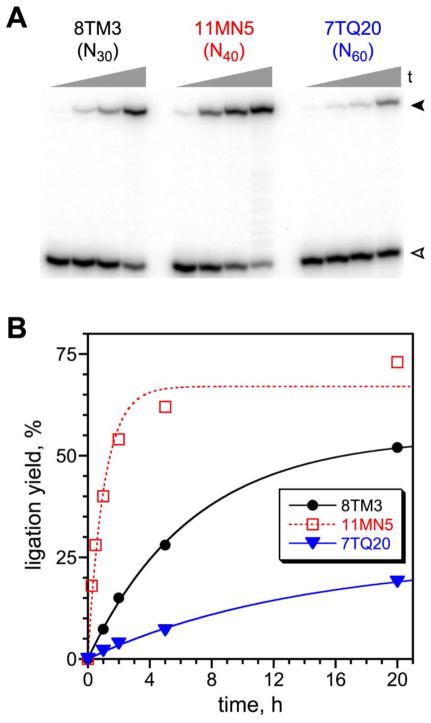
Functional nucleic acids are DNA and RNA aptamers that bind targets, or they are deoxyribozymes and ribozymes that have catalytic activity. These functional DNA and RNA sequences can be identified from random-sequence pools by in vitro selection, which requires choosing the length of the random region. Shorter random regions allow more complete coverage of sequence space but may not permit the structural complexity necessary for binding or catalysis. In contrast, longer random regions are sampled incompletely but may allow adoption of more complicated structures that enable function. In this study, we systematically examined random region length (N(20) through N(60)) for two particular deoxyribozyme catalytic activities, DNA cleavage and tyrosine-RNA nucleopeptide linkage formation. For both activities, we previously identified deoxyribozymes using only N(40) regions. In the case of DNA cleavage, here we found that shorter N(20) and N(30) regions allowed robust catalytic function, either by DNA hydrolysis or by DNA deglycosylation and strand scission via β-elimination, whereas longer N(50) and N(60) regions did not lead to catalytically active DNA sequences. Follow-up selections with N(20), N(30), and N(40) regions revealed an interesting interplay of metal ion cofactors and random region length. Separately, for Tyr-RNA linkage formation, N(30) and N(60) regions provided catalytically active sequences, whereas N(20) was unsuccessful, and the N(40) deoxyribozymes were functionally superior (in terms of rate and yield) to N(30) and N(60). Collectively, the results indicate that with future in vitro selection experiments for DNA and RNA catalysts, and by extension for aptamers, random region length should be an important experimental variable.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Doudna JA, Cech TR. The chemical repertoire of natural ribozymes. Nature. 2002;418:222–228. - PubMed
-
- Leung EK, Suslov N, Tuttle N, Sengupta R, Piccirilli JA. The mechanism of peptidyl transfer catalysis by the ribosome. Annu Rev Biochem. 2011;80:527–555. - PubMed
-
- Tuerk C, Gold L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science. 1990;249:505–510. - PubMed
-
- Ellington AD, Szostak JW. In vitro selection of RNA molecules that bind specific ligands. Nature. 1990;346:818–822. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
