

Covalent intermediate in the catalytic mechanism of the radical S-adenosyl-L-methionine methyl synthase RlmN trapped by mutagenesis
- PMID: 23088750
- PMCID: PMC3499099
- DOI: 10.1021/ja307855d
Covalent intermediate in the catalytic mechanism of the radical S-adenosyl-L-methionine methyl synthase RlmN trapped by mutagenesis
Abstract
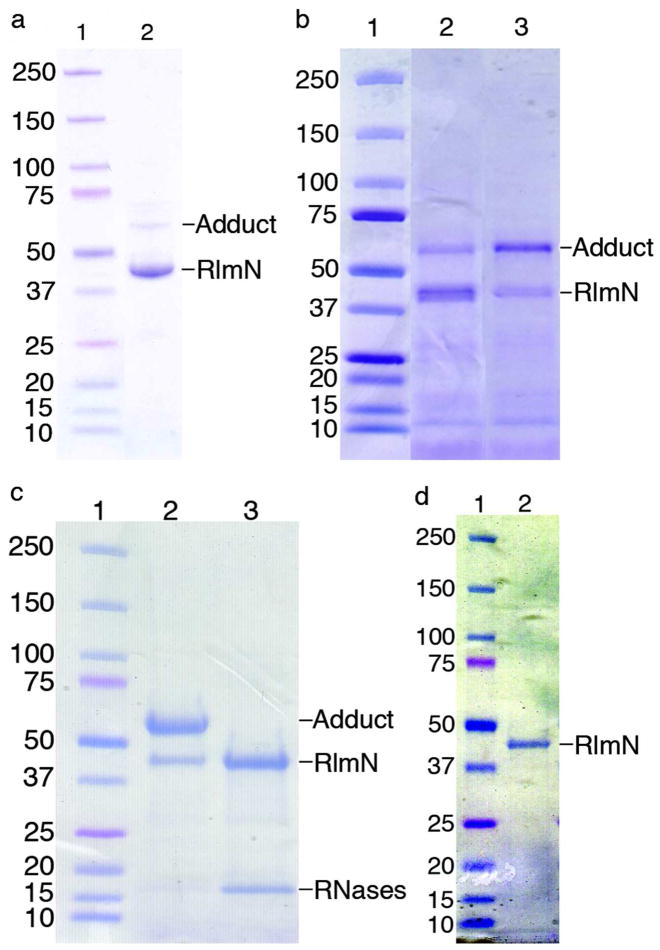
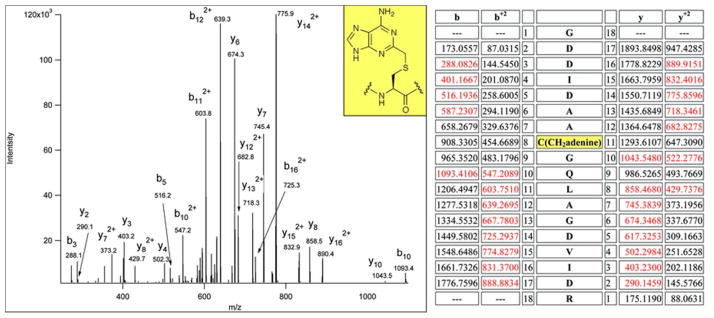
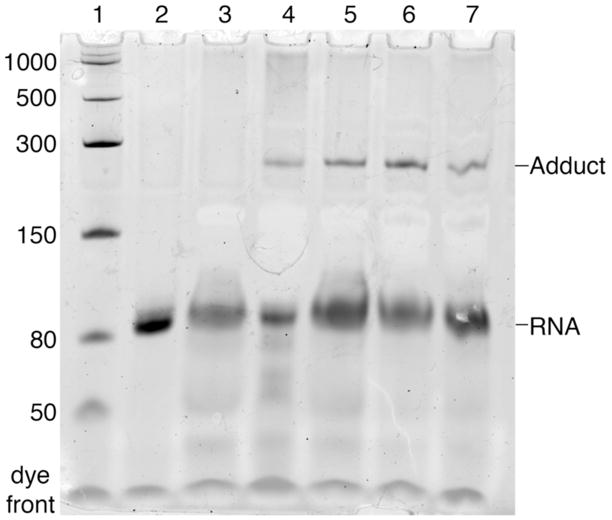
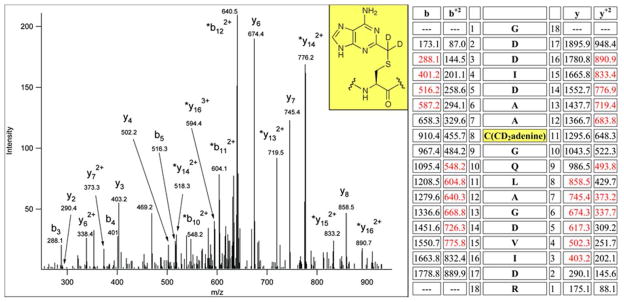
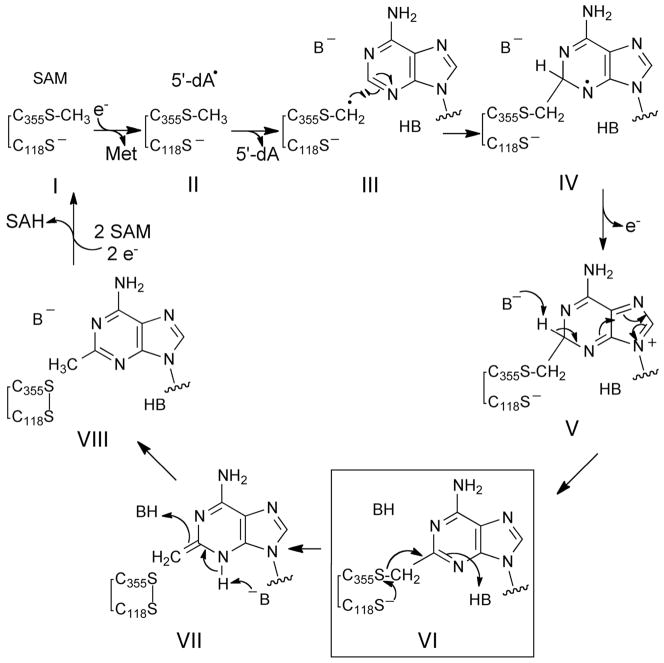
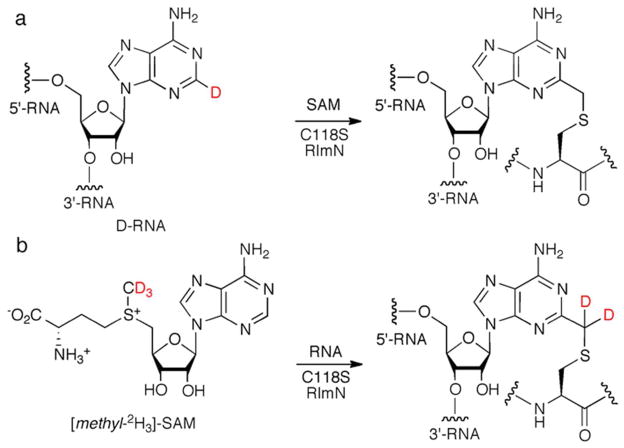
The posttranscriptional modification of ribosomal RNA (rRNA) modulates ribosomal function and confers resistance to antibiotics targeted to the ribosome. The radical S-adenosyl-L-methionine (SAM) methyl synthases, RlmN and Cfr, both methylate A2503 within the peptidyl transferase center of prokaryotic ribosomes, yielding 2-methyl- and 8-methyl-adenosine, respectively. The C2 and C8 positions of adenosine are unusual methylation substrates due to their electrophilicity. To accomplish this reaction, RlmN and Cfr use a shared radical-mediated mechanism. In addition to the radical SAM CX(3)CX(2)C motif, both RlmN and Cfr contain two conserved cysteine residues required for in vivo function, putatively to form (cysteine 355 in RlmN) and resolve (cysteine 118 in RlmN) a covalent intermediate needed to achieve this challenging transformation. Currently, there is no direct evidence for this proposed covalent intermediate. We have further investigated the roles of these conserved cysteines in the mechanism of RlmN. Cysteine 118 mutants of RlmN are unable to resolve the covalent intermediate, either in vivo or in vitro, enabling us to isolate and characterize this intermediate. Additionally, tandem mass spectrometric analyses of mutant RlmN reveal a methylene-linked adenosine modification at cysteine 355. Employing deuterium-labeled SAM and RNA substrates in vitro has allowed us to further clarify the mechanism of formation of this intermediate. Together, these experiments provide compelling evidence for the formation of a covalent intermediate species between RlmN and its rRNA substrate and well as the roles of the conserved cysteine residues in catalysis.
Figures
Similar articles
-
RlmN and Cfr are radical SAM enzymes involved in methylation of ribosomal RNA.J Am Chem Soc. 2010 Mar 24;132(11):3953-64. doi: 10.1021/ja910850y. J Am Chem Soc. 2010. PMID: 20184321 Free PMC article.
-
RNA methylation by radical SAM enzymes RlmN and Cfr proceeds via methylene transfer and hydride shift.Proc Natl Acad Sci U S A. 2011 Mar 8;108(10):3930-4. doi: 10.1073/pnas.1017781108. Epub 2011 Feb 22. Proc Natl Acad Sci U S A. 2011. PMID: 21368151 Free PMC article.
-
Structural basis for methyl transfer by a radical SAM enzyme.Science. 2011 May 27;332(6033):1089-92. doi: 10.1126/science.1205358. Epub 2011 Apr 28. Science. 2011. PMID: 21527678 Free PMC article.
-
Radical-mediated enzymatic methylation: a tale of two SAMS.Acc Chem Res. 2012 Apr 17;45(4):555-64. doi: 10.1021/ar200202c. Epub 2011 Nov 18. Acc Chem Res. 2012. PMID: 22097883 Free PMC article. Review.
-
Radical SAM-mediated methylation reactions.Curr Opin Chem Biol. 2013 Aug;17(4):597-604. doi: 10.1016/j.cbpa.2013.05.032. Epub 2013 Jul 5. Curr Opin Chem Biol. 2013. PMID: 23835516 Free PMC article. Review.
Cited by
-
Structure and Catalytic Mechanism of Radical SAM Methylases.Life (Basel). 2022 Oct 28;12(11):1732. doi: 10.3390/life12111732. Life (Basel). 2022. PMID: 36362886 Free PMC article. Review.
-
C2-methyladenosine in tRNA promotes protein translation by facilitating the decoding of tandem m2A-tRNA-dependent codons.Nat Commun. 2024 Feb 3;15(1):1025. doi: 10.1038/s41467-024-45166-6. Nat Commun. 2024. PMID: 38310199 Free PMC article.
-
Mutations in RNA methylating enzymes in disease.Curr Opin Chem Biol. 2017 Dec;41:20-27. doi: 10.1016/j.cbpa.2017.10.002. Epub 2017 Oct 20. Curr Opin Chem Biol. 2017. PMID: 29059606 Free PMC article. Review.
-
Mechanistic diversity of radical S-adenosylmethionine (SAM)-dependent methylation.J Biol Chem. 2015 Feb 13;290(7):3995-4002. doi: 10.1074/jbc.R114.607044. Epub 2014 Dec 4. J Biol Chem. 2015. PMID: 25477520 Free PMC article. Review.
-
Antibiotic resistance evolved via inactivation of a ribosomal RNA methylating enzyme.Nucleic Acids Res. 2016 Oct 14;44(18):8897-8907. doi: 10.1093/nar/gkw699. Epub 2016 Aug 5. Nucleic Acids Res. 2016. PMID: 27496281 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
