

Action of gibberellins on growth and metabolism of Arabidopsis plants associated with high concentration of carbon dioxide
- PMID: 23090585
- PMCID: PMC3510110
- DOI: 10.1104/pp.112.204842
Action of gibberellins on growth and metabolism of Arabidopsis plants associated with high concentration of carbon dioxide
Abstract
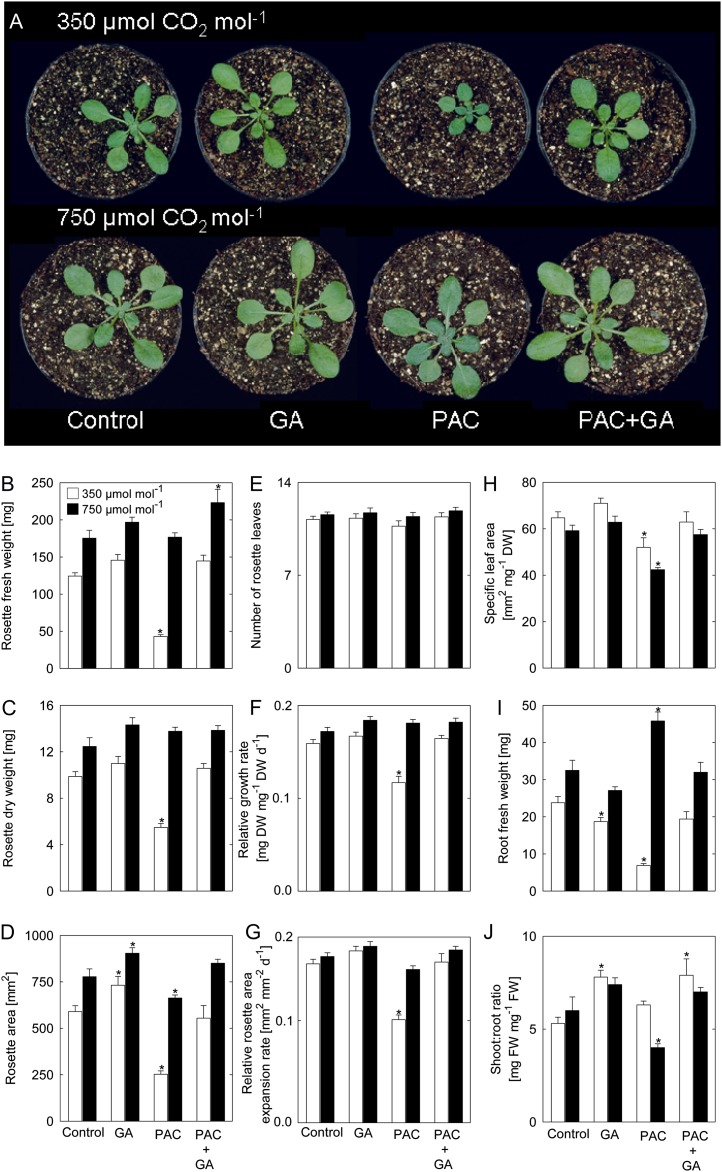
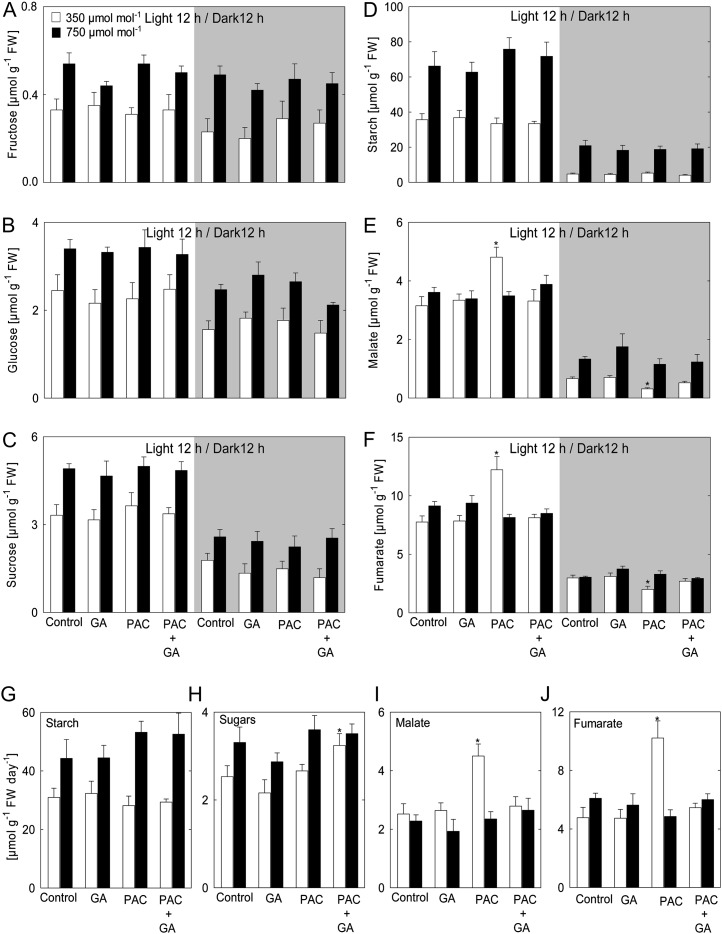
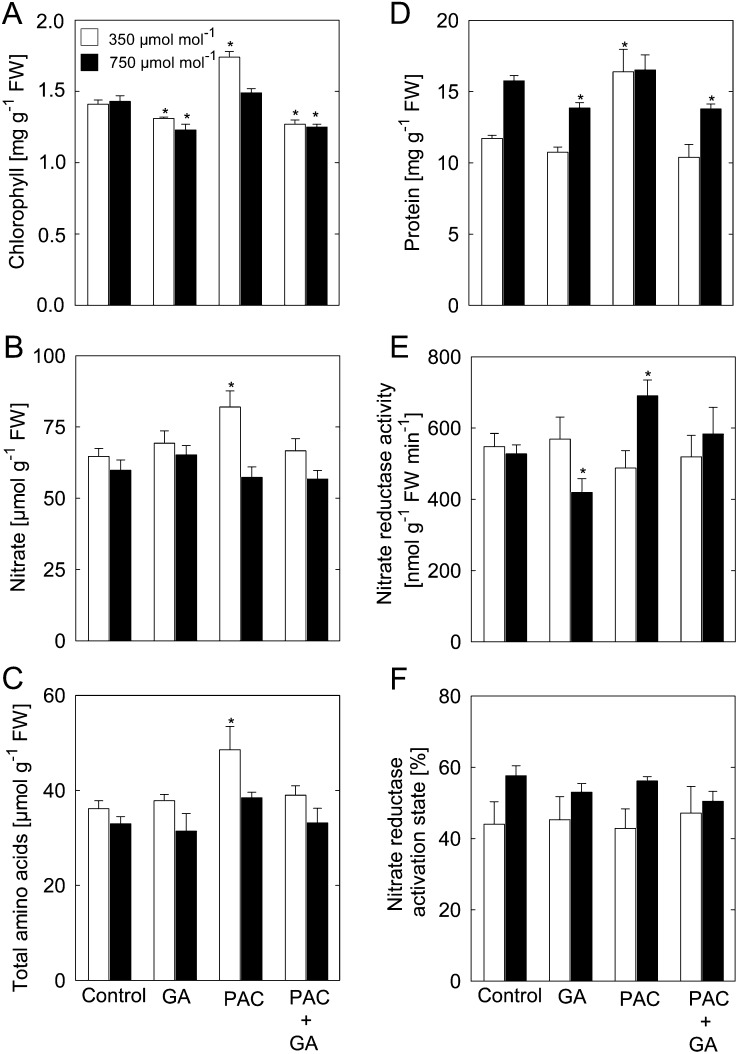
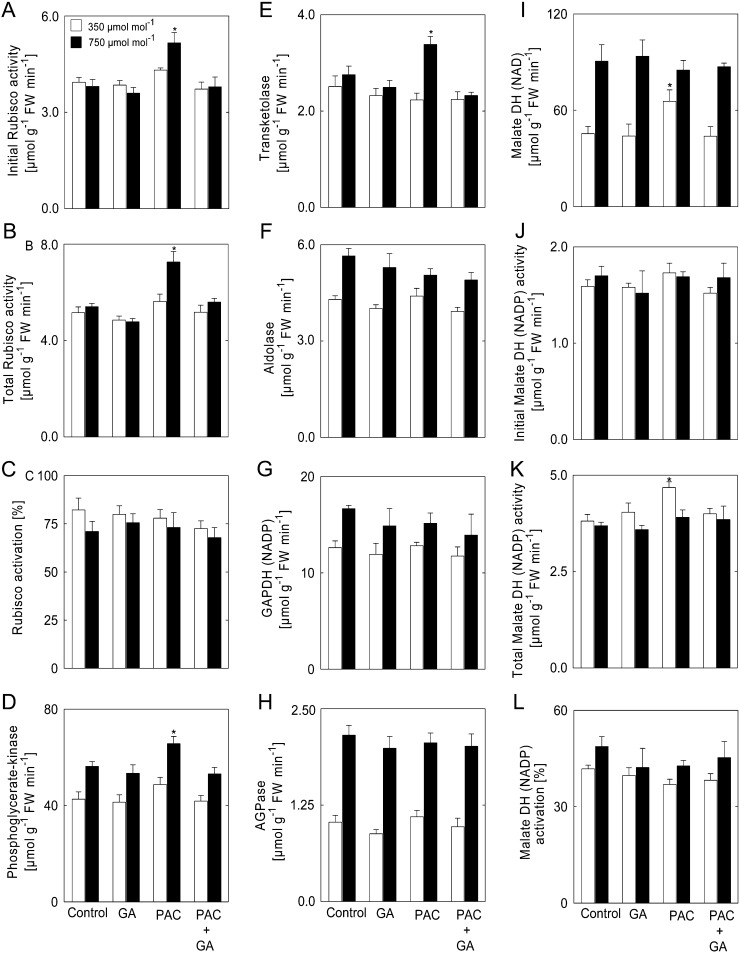
Although the positive effect of elevated CO(2) concentration [CO(2)] on plant growth is well known, it remains unclear whether global climate change will positively or negatively affect crop yields. In particular, relatively little is known about the role of hormone pathways in controlling the growth responses to elevated [CO(2)]. Here, we studied the impact of elevated [CO(2)] on plant biomass and metabolism in Arabidopsis (Arabidopsis thaliana) in relation to the availability of gibberellins (GAs). Inhibition of growth by the GA biosynthesis inhibitor paclobutrazol (PAC) at ambient [CO(2)] (350 µmol CO(2) mol(-1)) was reverted by elevated [CO(2)] (750 µmol CO(2) mol(-1)). Thus, we investigated the metabolic adjustment and modulation of gene expression in response to changes in growth of plants imposed by varying the GA regime in ambient and elevated [CO(2)]. In the presence of PAC (low-GA regime), the activities of enzymes involved in photosynthesis and inorganic nitrogen assimilation were markedly increased at elevated [CO(2)], whereas the activities of enzymes of organic acid metabolism were decreased. Under ambient [CO(2)], nitrate, amino acids, and protein accumulated upon PAC treatment; however, this was not the case when plants were grown at elevated [CO(2)]. These results suggest that only under ambient [CO(2)] is GA required for the integration of carbohydrate and nitrogen metabolism underlying optimal biomass determination. Our results have implications concerning the action of the Green Revolution genes in future environmental conditions.
Figures
References
-
- Ainsworth EA. (2008) Rice production in a changing climate: a meta-analysis of responses to elevated carbon dioxide and elevated ozone concentrations. Glob Change Biol 14: 1642–1650
-
- Ainsworth EA, Davey PA, Bernacchi CJ, Dermody OC, Heaton EA, Moore DJ, Morgan PB, Naidu SL, Ra HSY, Zhu XG, et al. (2002) A meta-analysis of elevated [CO2] effects on soybean (Glycine max) physiology, growth and yield. Glob Change Biol 8: 695–709
-
- Araújo WL, Tohge T, Osorio S, Lohse M, Balbo I, Krahnert I, Sienkiewicz-Porzucek A, Usadel B, Nunes-Nesi A, Fernie AR. (2012) Antisense inhibition of the 2-oxoglutarate dehydrogenase complex in tomato demonstrates its importance for plant respiration and during leaf senescence and fruit maturation. Plant Cell 24: 2328–2351 - PMC - PubMed
-
- Bae H, Sicher R. (2004) Changes of soluble protein expression and leaf metabolite levels in Arabidopsis thaliana grown in elevated atmospheric carbon dioxide. Field Crops Res 90: 61–73
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
