

Uncoupling of nucleotide hydrolysis and polymerization in the ParA protein superfamily disrupts DNA segregation dynamics
- PMID: 23093445
- PMCID: PMC3522256
- DOI: 10.1074/jbc.M112.410324
Uncoupling of nucleotide hydrolysis and polymerization in the ParA protein superfamily disrupts DNA segregation dynamics
Abstract
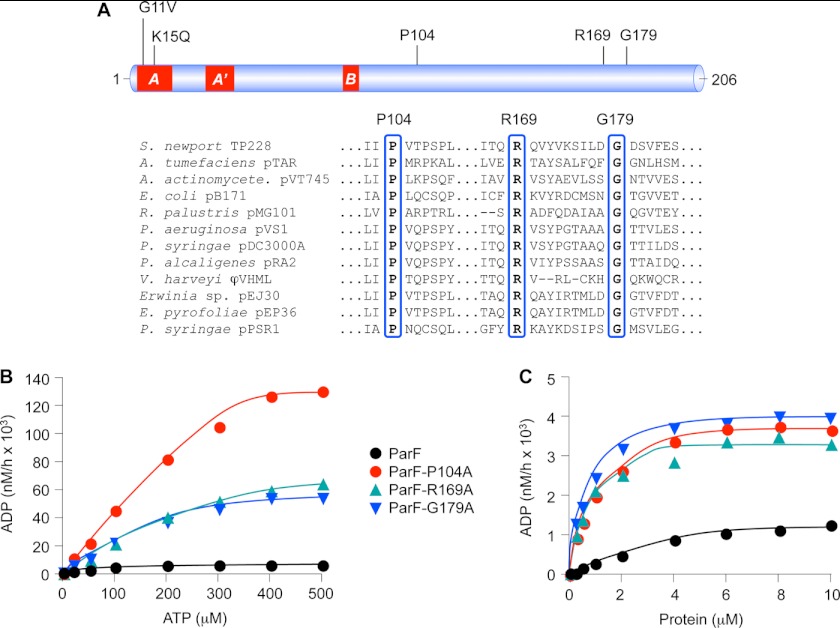
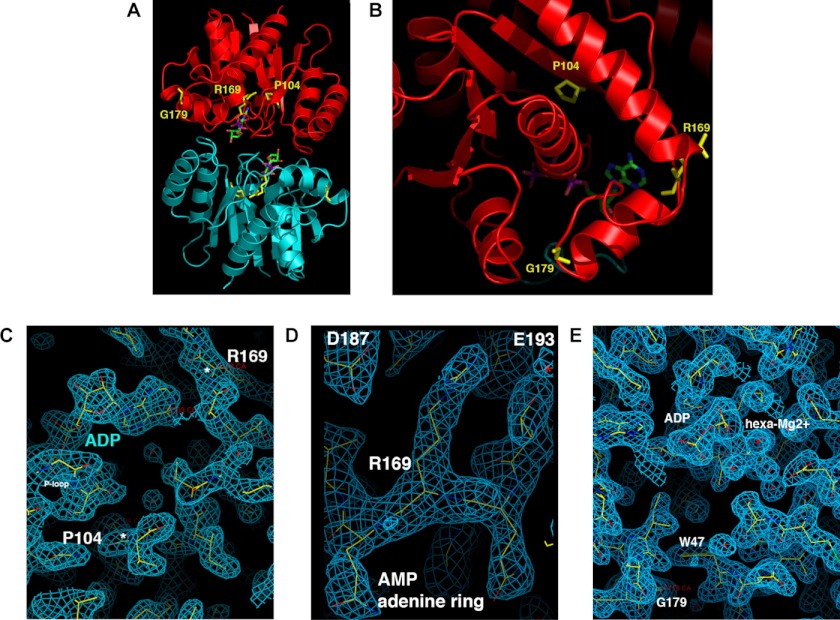
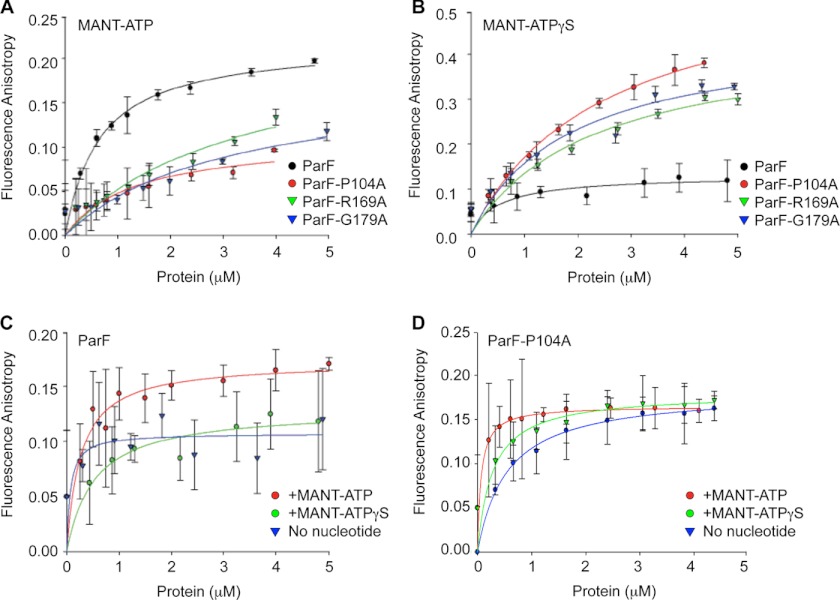
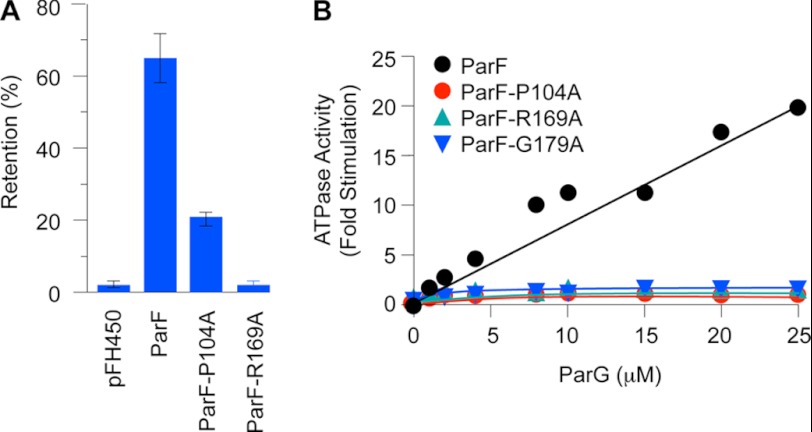
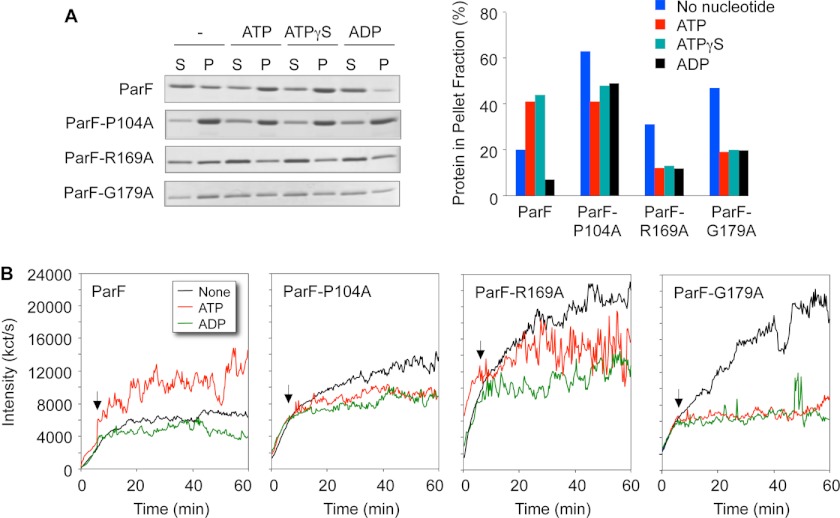
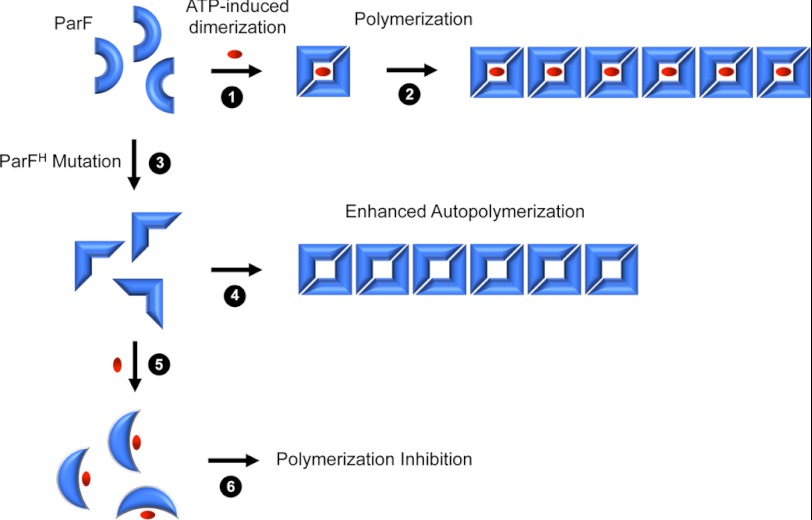
DNA segregation in bacteria is mediated most frequently by proteins of the ParA superfamily that transport DNA molecules attached via the segrosome nucleoprotein complex. Segregation is governed by a cycle of ATP-induced polymerization and subsequent depolymerization of the ParA factor. Here, we establish that hyperactive ATPase variants of the ParA homolog ParF display altered segrosome dynamics that block accurate DNA segregation. An arginine finger-like motif in the ParG centromere-binding factor augments ParF ATPase activity but is ineffective in stimulating nucleotide hydrolysis by the hyperactive proteins. Moreover, whereas polymerization of wild-type ParF is accelerated by ATP and inhibited by ADP, filamentation of the mutated proteins is blocked indiscriminately by nucleotides. The mutations affect a triplet of conserved residues that are situated neither in canonical nucleotide binding and hydrolysis motifs in the ParF tertiary structure nor at interfaces implicated in ParF polymerization. Instead the residues are involved in shaping the contours of the binding pocket so that nucleotide binding locks the mutant proteins into a configuration that is refractory to polymerization. Thus, the architecture of the pocket not only is crucial for optimal ATPase kinetics but also plays a key role in the polymerization dynamics of ParA proteins that drive DNA segregation ubiquitously in procaryotes.
Figures
Similar articles
-
The tail of the ParG DNA segregation protein remodels ParF polymers and enhances ATP hydrolysis via an arginine finger-like motif.Proc Natl Acad Sci U S A. 2007 Feb 6;104(6):1811-6. doi: 10.1073/pnas.0607216104. Epub 2007 Jan 29. Proc Natl Acad Sci U S A. 2007. PMID: 17261809 Free PMC article.
-
Promiscuous stimulation of ParF protein polymerization by heterogeneous centromere binding factors.J Mol Biol. 2007 Nov 16;374(1):1-8. doi: 10.1016/j.jmb.2007.09.025. Epub 2007 Sep 14. J Mol Biol. 2007. PMID: 17920627
-
Bacterial DNA segregation dynamics mediated by the polymerizing protein ParF.EMBO J. 2005 Apr 6;24(7):1453-64. doi: 10.1038/sj.emboj.7600619. Epub 2005 Mar 10. EMBO J. 2005. PMID: 15775965 Free PMC article.
-
Structural mechanism of ATP-induced polymerization of the partition factor ParF: implications for DNA segregation.J Biol Chem. 2012 Jul 27;287(31):26146-54. doi: 10.1074/jbc.M112.373696. Epub 2012 Jun 6. J Biol Chem. 2012. PMID: 22674577 Free PMC article.
-
The bacterial segrosome: a dynamic nucleoprotein machine for DNA trafficking and segregation.Nat Rev Microbiol. 2006 Feb;4(2):133-43. doi: 10.1038/nrmicro1342. Nat Rev Microbiol. 2006. PMID: 16415929 Review.
Cited by
-
Maintenance of multipartite genome system and its functional significance in bacteria.J Genet. 2018 Sep;97(4):1013-1038. J Genet. 2018. PMID: 30262715 Review.
-
Controlling the Revolving and Rotating Motion Direction of Asymmetric Hexameric Nanomotor by Arginine Finger and Channel Chirality.ACS Nano. 2019 Jun 25;13(6):6207-6223. doi: 10.1021/acsnano.8b08849. Epub 2019 May 28. ACS Nano. 2019. PMID: 31067030 Free PMC article. Review.
-
Mutational analysis of the F plasmid partitioning protein ParA reveals residues required for oligomerization and plasmid maintenance.Nucleic Acids Res. 2025 Jun 20;53(12):gkaf537. doi: 10.1093/nar/gkaf537. Nucleic Acids Res. 2025. PMID: 40568943 Free PMC article.
-
Genome Segregation by the Venus Flytrap Mechanism: Probing the Interaction Between the ParF ATPase and the ParG Centromere Binding Protein.Front Mol Biosci. 2020 Jun 16;7:108. doi: 10.3389/fmolb.2020.00108. eCollection 2020. Front Mol Biosci. 2020. PMID: 32613008 Free PMC article.
-
Catching a Walker in the Act-DNA Partitioning by ParA Family of Proteins.Front Microbiol. 2022 May 26;13:856547. doi: 10.3389/fmicb.2022.856547. eCollection 2022. Front Microbiol. 2022. PMID: 35694299 Free PMC article. Review.
References
-
- Anand S. P., Akhtar P., Tinsley E., Watkins S. C., Khan S. A. (2008) GTP-dependent polymerization of the tubulin-like RepX replication protein encoded by the pXO1 plasmid of Bacillus anthracis. Mol. Microbiol. 67, 881–890 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- G0801162/MRC_/Medical Research Council/United Kingdom
- R01 GM074815/GM/NIGMS NIH HHS/United States
- BB/G003114/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G003114/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- GM074815/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
