

Endothelial deletion of murine Jag1 leads to valve calcification and congenital heart defects associated with Alagille syndrome
- PMID: 23095891
- PMCID: PMC3509736
- DOI: 10.1242/dev.084871
Endothelial deletion of murine Jag1 leads to valve calcification and congenital heart defects associated with Alagille syndrome
Abstract
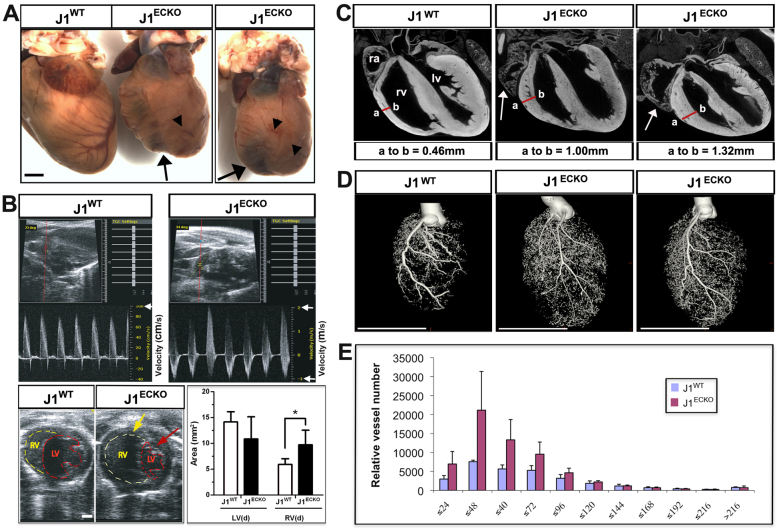
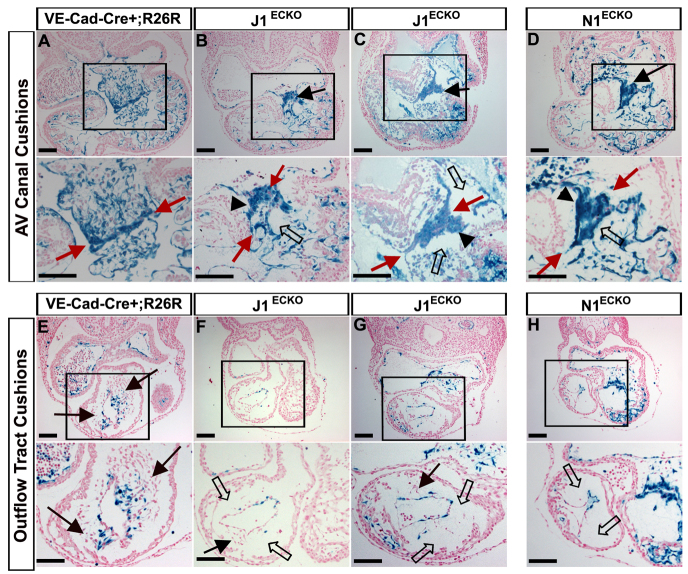
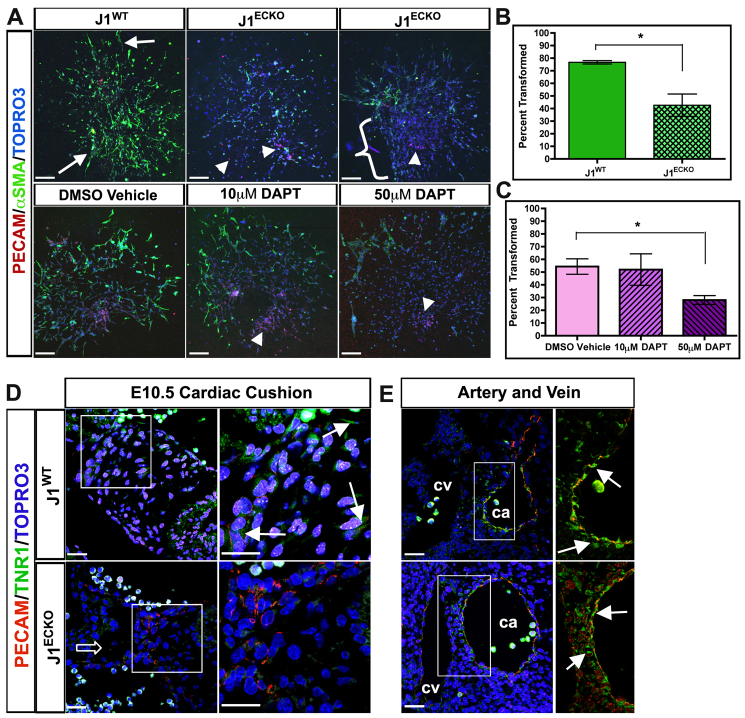
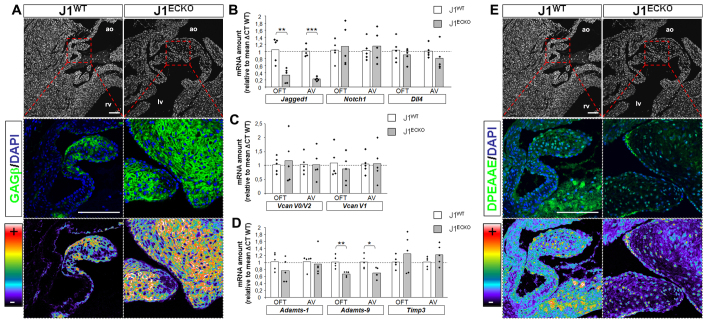
The Notch signaling pathway is an important contributor to the development and homeostasis of the cardiovascular system. Not surprisingly, mutations in Notch receptors and ligands have been linked to a variety of hereditary diseases that impact both the heart and the vasculature. In particular, mutations in the gene encoding the human Notch ligand jagged 1 result in a multisystem autosomal dominant disorder called Alagille syndrome, which includes tetralogy of Fallot among its more severe cardiac pathologies. Jagged 1 is expressed throughout the developing embryo, particularly in endothelial cells. Here, we demonstrate that endothelial-specific deletion of Jag1 leads to cardiovascular defects in both embryonic and adult mice that are reminiscent of those in Alagille syndrome. Mutant mice display right ventricular hypertrophy, overriding aorta, ventricular septal defects, coronary vessel abnormalities and valve defects. Examination of mid-gestational embryos revealed that the loss of Jag1, similar to the loss of Notch1, disrupts endothelial-to-mesenchymal transition during endocardial cushion formation. Furthermore, adult mutant mice exhibit cardiac valve calcifications associated with abnormal matrix remodeling and induction of bone morphogenesis. This work shows that the endothelium is responsible for the wide spectrum of cardiac phenotypes displayed in Alagille Syndrome and it demonstrates a crucial role for Jag1 in valve morphogenesis.
Figures




Similar articles
-
A mouse model of Alagille syndrome: Notch2 as a genetic modifier of Jag1 haploinsufficiency.Development. 2002 Feb;129(4):1075-82. doi: 10.1242/dev.129.4.1075. Development. 2002. PMID: 11861489
-
Functional role of Notch signaling in the developing and postnatal heart.J Mol Cell Cardiol. 2008 Oct;45(4):495-504. doi: 10.1016/j.yjmcc.2008.02.273. Epub 2008 Mar 10. J Mol Cell Cardiol. 2008. PMID: 18410944 Review.
-
Variable expression of Alagille syndrome in a family with a new JAG1 gene mutation.Cardiol Young. 2016 Jan;26(1):164-7. doi: 10.1017/S1047951114002753. Epub 2015 Jan 23. Cardiol Young. 2016. PMID: 25613755
-
Tetralogy of fallot and other congenital heart defects in Hey2 mutant mice.Curr Biol. 2002 Sep 17;12(18):1605-10. doi: 10.1016/s0960-9822(02)01149-1. Curr Biol. 2002. PMID: 12372254
-
Jagged1 (JAG1): Structure, expression, and disease associations.Gene. 2016 Jan 15;576(1 Pt 3):381-4. doi: 10.1016/j.gene.2015.10.065. Epub 2015 Nov 6. Gene. 2016. PMID: 26548814 Free PMC article. Review.
Cited by
-
Endothelial Heterogeneity in Development and Wound Healing.Cells. 2021 Sep 7;10(9):2338. doi: 10.3390/cells10092338. Cells. 2021. PMID: 34571987 Free PMC article. Review.
-
Endocardial Brg1 disruption illustrates the developmental origins of semilunar valve disease.Dev Biol. 2015 Nov 1;407(1):158-72. doi: 10.1016/j.ydbio.2015.06.015. Epub 2015 Jun 20. Dev Biol. 2015. PMID: 26100917 Free PMC article.
-
Jagged1 Instructs Macrophage Differentiation in Leprosy.PLoS Pathog. 2016 Aug 17;12(8):e1005808. doi: 10.1371/journal.ppat.1005808. eCollection 2016 Aug. PLoS Pathog. 2016. PMID: 27532668 Free PMC article.
-
Genetic basis of aortic valvular disease.Curr Opin Cardiol. 2017 May;32(3):239-245. doi: 10.1097/HCO.0000000000000384. Curr Opin Cardiol. 2017. PMID: 28157139 Free PMC article.
-
Foetal hepatic progenitor cells assume a cholangiocytic cell phenotype during two-dimensional pre-culture.Sci Rep. 2016 Jun 23;6:28283. doi: 10.1038/srep28283. Sci Rep. 2016. PMID: 27335264 Free PMC article.
References
-
- Alva J. A., Zovein A. C., Monvoisin A., Murphy T., Salazar A., Harvey N. L., Carmeliet P., Iruela-Arispe M. L. (2006). VE-Cadherin-Cre-recombinase transgenic mouse: a tool for lineage analysis and gene deletion in endothelial cells. Dev. Dyn. 235, 759-767 - PubMed
-
- Benedito R., Roca C., Sörensen I., Adams S., Gossler A., Fruttiger M., Adams R. H. (2009). The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell 137, 1124-1135 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
