

Influenza A viruses grow in human pancreatic cells and cause pancreatitis and diabetes in an animal model
- PMID: 23097451
- PMCID: PMC3536404
- DOI: 10.1128/JVI.00714-12
Influenza A viruses grow in human pancreatic cells and cause pancreatitis and diabetes in an animal model
Abstract
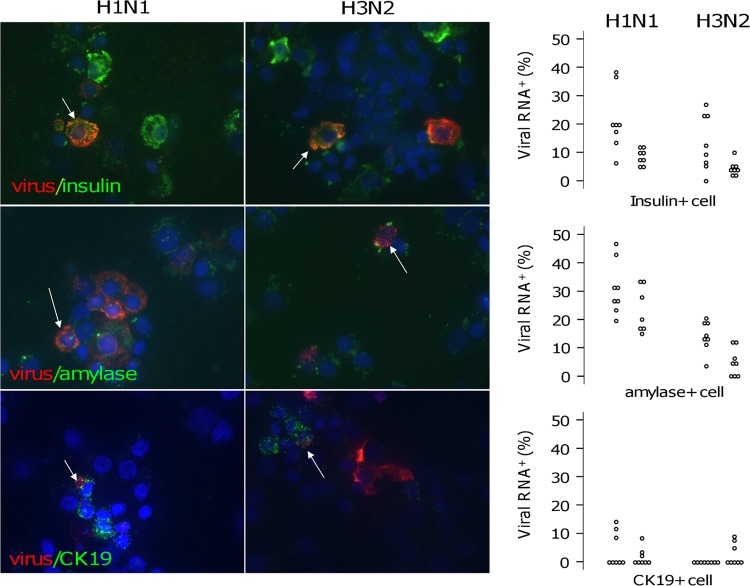
Influenza A viruses commonly cause pancreatitis in naturally and experimentally infected animals. In this study, we report the results of in vivo investigations carried out to establish whether influenza virus infection could cause metabolic disorders linked to pancreatic infection. In addition, in vitro tests in human pancreatic islets and in human pancreatic cell lines were performed to evaluate viral growth and cell damage. Infection of an avian model with two low-pathogenicity avian influenza isolates caused pancreatic damage resulting in hyperlipasemia in over 50% of subjects, which evolved into hyperglycemia and subsequently diabetes. Histopathology of the pancreas showed signs of an acute infection resulting in severe fibrosis and disruption of the structure of the organ. Influenza virus nucleoprotein was detected by immunohistochemistry (IHC) in the acinar tissue. Human seasonal H1N1 and H3N2 viruses and avian H7N1 and H7N3 influenza virus isolates were able to infect a selection of human pancreatic cell lines. Human viruses were also shown to be able to infect human pancreatic islets. In situ hybridization assays indicated that viral nucleoprotein could be detected in beta cells. The cytokine activation profile indicated a significant increase of MIG/CXCL9, IP-10/CXCL10, RANTES/CCL5, MIP1b/CCL4, Groa/CXCL1, interleukin 8 (IL-8)/CXCL8, tumor necrosis factor alpha (TNF-α), and IL-6. Our findings indicate that influenza virus infection may play a role as a causative agent of pancreatitis and diabetes in humans and other mammals.
Figures








References
-
- Capua I, Alexander DJ. 2009. Avian influenza and Newcastle Disease, a field and laboratory manual. Springer, Berlin, Germany
-
- Harder TC, Vahlenkamp TW. 2010. Influenza virus infections in dogs and cats. Vet. Immunol. Immunopathol. 134:54–60 - PubMed
-
- Kuiken T, Rimmelzwaan G, van Riel D, van Amerongen G, Baars M, Fouchier R, Osterhaus A. 2004. Avian H5N1 influenza in cats. Science 306:241. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
