

Functional analysis of in-frame indel ARID1A mutations reveals new regulatory mechanisms of its tumor suppressor functions
- PMID: 23097632
- PMCID: PMC3479842
- DOI: 10.1593/neo.121218
Functional analysis of in-frame indel ARID1A mutations reveals new regulatory mechanisms of its tumor suppressor functions
Abstract
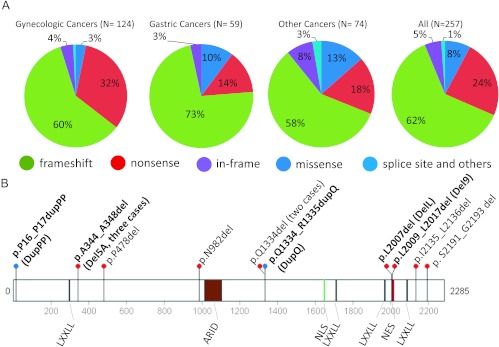
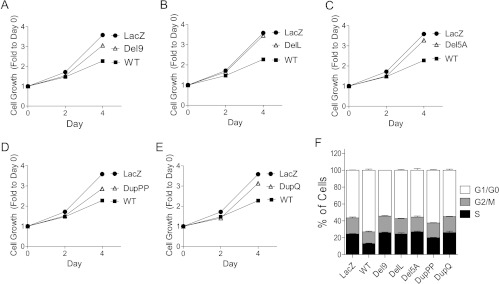
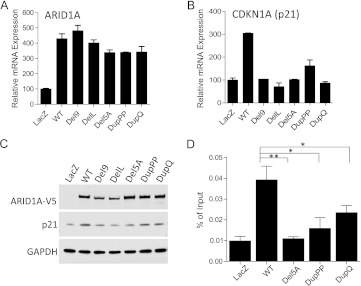
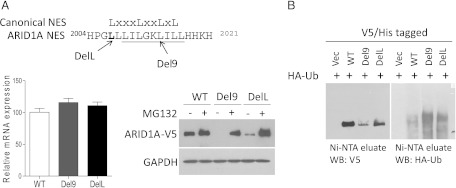
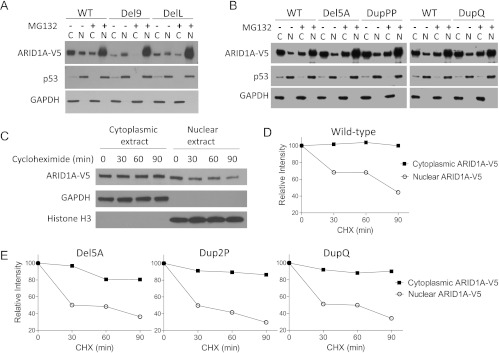
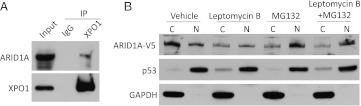
AT-rich interactive domain 1A (ARID1A) has emerged as a new tumor suppressor in which frequent somatic mutations have been identified in several types of human cancers. Although most ARID1A somatic mutations are frame-shift or nonsense mutations that contribute to mRNA decay and loss of protein expression, 5% of ARID1A mutations are in-frame insertions or deletions (indels) that involve only a small stretch of peptides. Naturally occurring in-frame indel mutations provide unique and useful models to explore the biology and regulatory role of ARID1A. In this study, we analyzed indel mutations identified in gynecological cancers to determine how these mutations affect the tumor suppressor function of ARID1A. Our results demonstrate that all in-frame mutants analyzed lost their ability to inhibit cellular proliferation or activate transcription of CDKN1A, which encodes p21, a downstream effector of ARID1A. We also showed that ARID1A is a nucleocytoplasmic protein whose stability depends on its subcellular localization. Nuclear ARID1A is less stable than cytoplasmic ARID1A because ARID1A is rapidly degraded by the ubiquitin-proteasome system in the nucleus. In-frame deletions affecting the consensus nuclear export signal reduce steady-state protein levels of ARID1A. This defect in nuclear exportation leads to nuclear retention and subsequent degradation. Our findings delineate a mechanism underlying the regulation of ARID1A subcellular distribution and protein stability and suggest that targeting the nuclear ubiquitin-proteasome system can increase the amount of the ARID1A protein in the nucleus and restore its tumor suppressor functions.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials