

Salubrinal protects against tunicamycin and hypoxia induced cardiomyocyte apoptosis via the PERK-eIF2α signaling pathway
- PMID: 23097656
- PMCID: PMC3470025
- DOI: 10.3724/SP.J.1263.2012.02292
Salubrinal protects against tunicamycin and hypoxia induced cardiomyocyte apoptosis via the PERK-eIF2α signaling pathway
Abstract
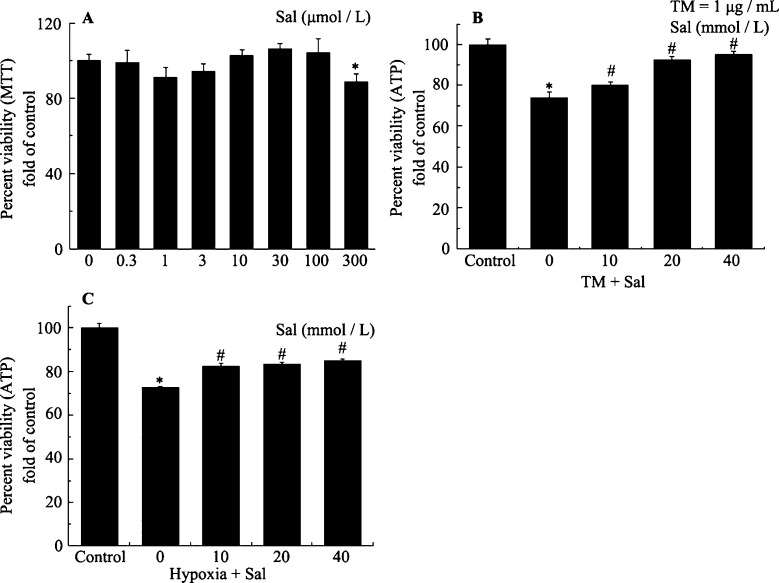
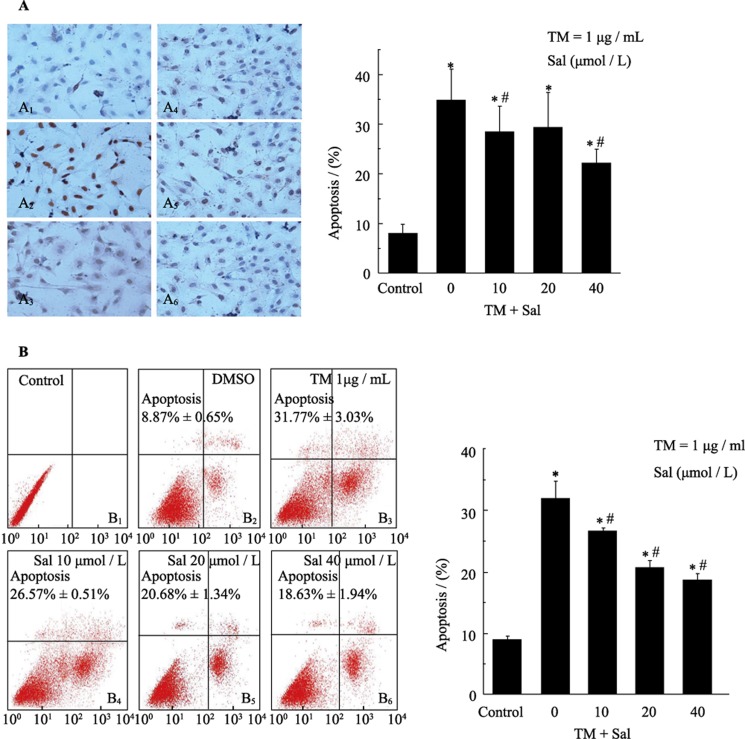
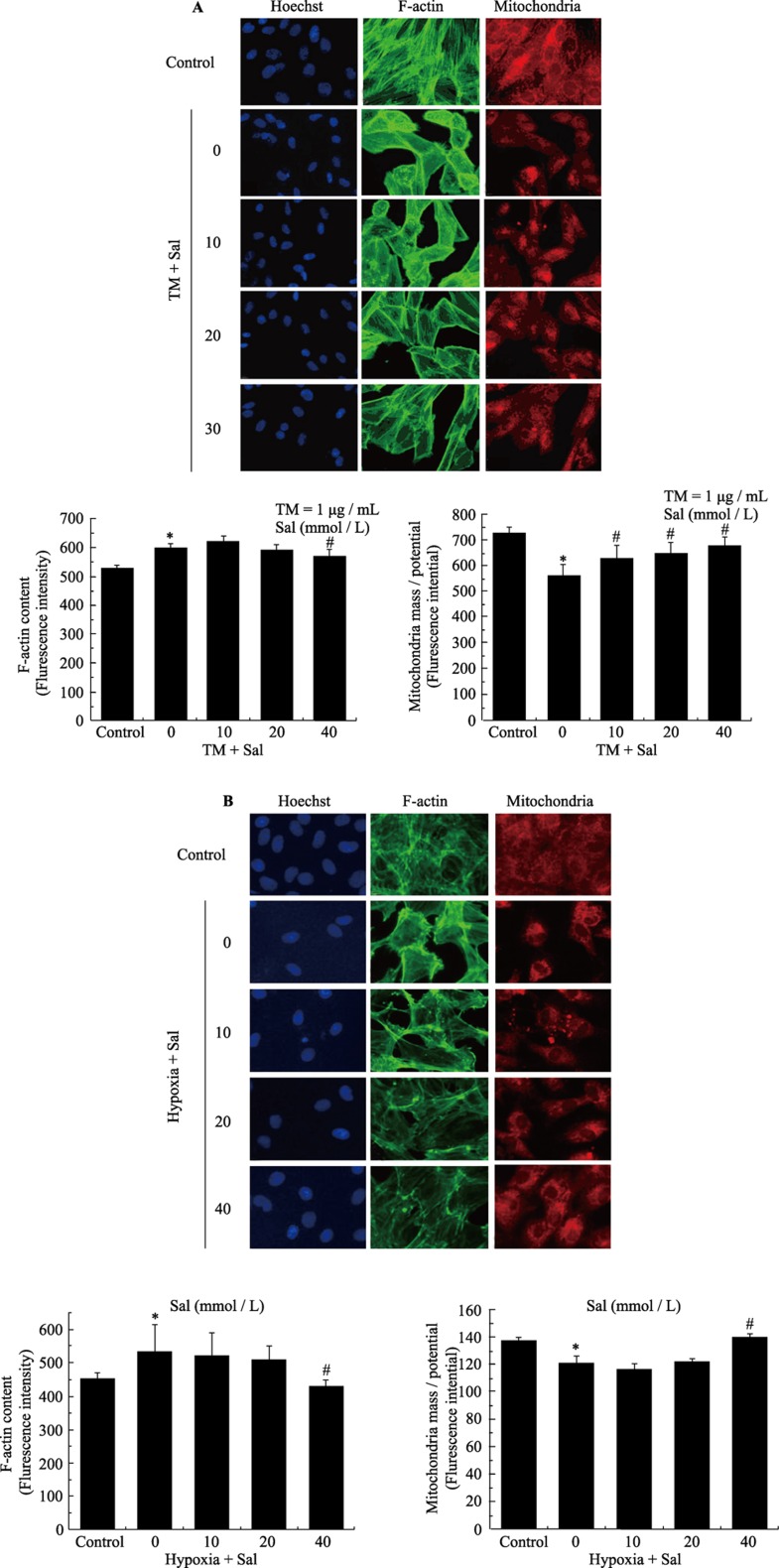
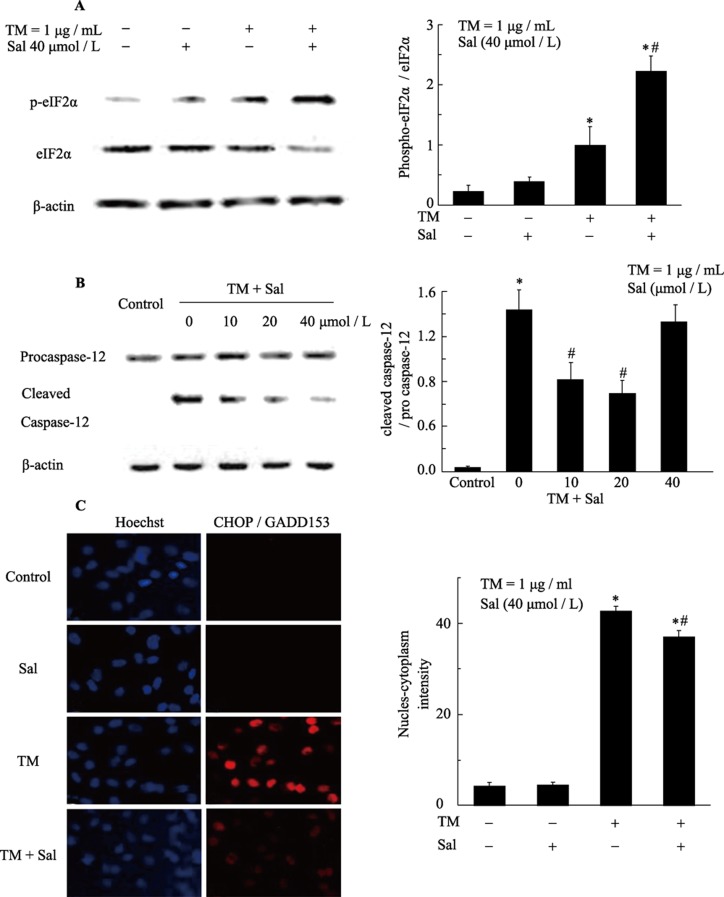
Objectives: This study examined the protective effect of salubrinal and the mechanism underlying this protection against tunicamycin (TM)- and hypoxia-induced apoptosis in rat cardiomyocytes.
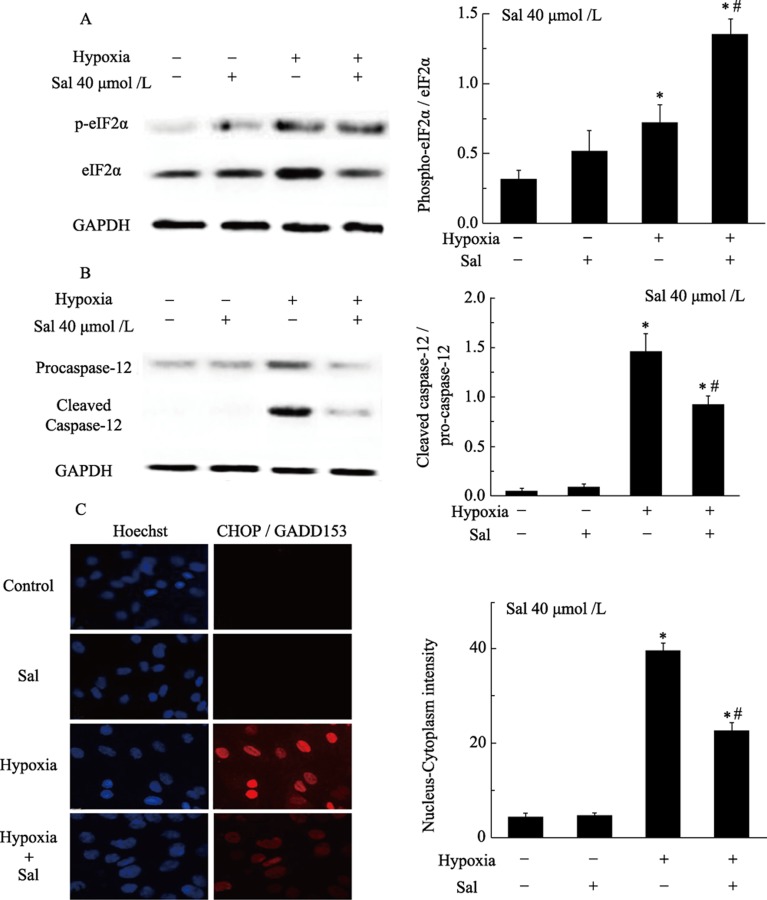
Methods: Neonatal rat cardiomyocytes were cultured from the ventricles of 1-day-old Wistar rats. Cells were exposed to different concentrations of salubrinal (10, 20, and 40 µmol/L) for 30 min followed by TM treatment or hypoxia for 36 h. Apoptosis was measured by a multiparameter HCS (high content screening) apoptosis assay, TUNEL assay and flow cytometry. The phosphorylation of eukaryotic translation initiation factor 2 subunit alpha (eIF2α) and the expression of cleaved caspase-12 were determined by Western blotting. C/EBP homologous protein (CHOP) was detected by immunocytochemistry.
Results: HCS, TUNEL assays and flow cytometry showed that salubrinal protected cardiomyocytes against apoptosis induced by TM or hypoxia. Western blotting showed that salubrinal protected cardiomyocytes against apoptosis by inducing eIF2α phosphorylation and down-regulating the expression of the endoplasmic reticulum stress-mediated apoptotic proteins, CHOP and cleaved caspase-12.
Conclusions: Our study suggests that salubrinal protects rat cardiomyocytes against TM- or hypoxia-associated apoptosis via a mechanism involving the inhibition of ER stress-mediated apoptosis.
Keywords: Apoptosis; Cell protection; Endoplasmic reticulum stress; Rat cardiomyocytes; Salubrinal.
Figures
References
-
- Tajiri S, Oyadomari S, Yano S, et al. Ischemia-induced neuronal cell death is mediated by the endoplasmic reticulum stress pathway involving CHOP. Cell Death Differ. 2004;11:403–415. - PubMed
-
- Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Genes Dev. 1999;13:1211–1233. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials