

ATP-binding cassette transporter G1 intrinsically regulates invariant NKT cell development
- PMID: 23100511
- PMCID: PMC3776580
- DOI: 10.4049/jimmunol.1201570
ATP-binding cassette transporter G1 intrinsically regulates invariant NKT cell development
Abstract
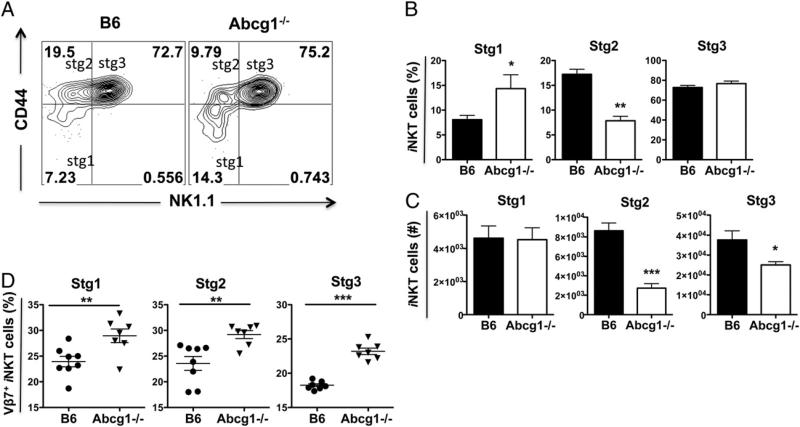
ATP-binding cassette transporter G1 (ABCG1) plays a role in the intracellular transport of cholesterol. Invariant NKT (iNKT) cells are a subpopulation of T lymphocytes that recognize glycolipid Ags. In this study, we demonstrate that ABCG1 regulates iNKT cell development and functions in a cell-intrinsic manner. Abcg1(-/-) mice displayed reduced frequencies of iNKT cells in thymus and periphery. Thymic iNKT cells deficient in ABCG1 had reduced membrane lipid raft content, and showed impaired proliferation and defective maturation during the early stages of development. Moreover, we found that Abcg1(-/-) mice possess a higher frequency of Vβ7(+) iNKT cells, suggesting alterations in iNKT cell thymic selection. Furthermore, in response to CD3ε/CD28 stimulation, Abcg1(-/-) thymic iNKT cells showed reduced production of IL-4 but increased production of IFN-γ. Our results demonstrate that changes in intracellular cholesterol homeostasis by ABCG1 profoundly impact iNKT cell development and function.
Figures





Similar articles
-
ATP Binding Cassette Transporter ABCA7 Regulates NKT Cell Development and Function by Controlling CD1d Expression and Lipid Raft Content.Sci Rep. 2017 Jan 16;7:40273. doi: 10.1038/srep40273. Sci Rep. 2017. PMID: 28091533 Free PMC article.
-
Critical role for invariant chain in CD1d-mediated selection and maturation of Vα14-invariant NKT cells.Immunol Lett. 2011 Sep 30;139(1-2):33-41. doi: 10.1016/j.imlet.2011.04.012. Epub 2011 May 5. Immunol Lett. 2011. PMID: 21565221 Free PMC article.
-
ATP-binding cassette transporter G1 negatively regulates thymocyte and peripheral lymphocyte proliferation.J Immunol. 2010 Jan 1;184(1):173-83. doi: 10.4049/jimmunol.0902372. Epub 2009 Nov 30. J Immunol. 2010. PMID: 19949102 Free PMC article.
-
The Role of Autophagy in iNKT Cell Development.Front Immunol. 2018 Nov 14;9:2653. doi: 10.3389/fimmu.2018.02653. eCollection 2018. Front Immunol. 2018. PMID: 30487800 Free PMC article. Review.
-
The ins and outs of type I iNKT cell development.Mol Immunol. 2019 Jan;105:116-130. doi: 10.1016/j.molimm.2018.09.023. Epub 2018 Nov 28. Mol Immunol. 2019. PMID: 30502719 Free PMC article. Review.
Cited by
-
Unveiling tumor immune evasion mechanisms: abnormal expression of transporters on immune cells in the tumor microenvironment.Front Immunol. 2023 Jul 21;14:1225948. doi: 10.3389/fimmu.2023.1225948. eCollection 2023. Front Immunol. 2023. PMID: 37545500 Free PMC article. Review.
-
Specific deletion of LDL receptor-related protein on macrophages has skewed in vivo effects on cytokine production by invariant natural killer T cells.PLoS One. 2014 Jul 22;9(7):e102236. doi: 10.1371/journal.pone.0102236. eCollection 2014. PLoS One. 2014. PMID: 25050824 Free PMC article.
-
Hypercholesterolemia Increases Colorectal Cancer Incidence by Reducing Production of NKT and γδ T Cells from Hematopoietic Stem Cells.Cancer Res. 2017 May 1;77(9):2351-2362. doi: 10.1158/0008-5472.CAN-16-1916. Epub 2017 Mar 1. Cancer Res. 2017. PMID: 28249902 Free PMC article.
-
Regulation of Adaptive Immunity in Health and Disease by Cholesterol Metabolism.Curr Allergy Asthma Rep. 2015 Aug;15(8):48. doi: 10.1007/s11882-015-0548-7. Curr Allergy Asthma Rep. 2015. PMID: 26149587 Free PMC article. Review.
-
ATP-dependent transporters: emerging players at the crossroads of immunity and metabolism.Front Immunol. 2023 Oct 31;14:1286696. doi: 10.3389/fimmu.2023.1286696. eCollection 2023. Front Immunol. 2023. PMID: 38022644 Free PMC article. Review.
References
-
- Godfrey DI, MacDonald HR, Kronenberg M, Smyth MJ, Van Kaer L. NKT cells: what's in a name? Nat. Rev. Immunol. 2004;4:231–237. - PubMed
-
- Kinjo Y, Wu D, Kim G, Xing GW, Poles MA, Ho DD, Tsuji M, Kawahara K, Wong CH, Kronenberg M. Recognition of bacterial glycosphingolipids by natural killer T cells. Nature. 2005;434:520–525. - PubMed
-
- Mattner J, Debord KL, Ismail N, Goff RD, Cantu C, III, Zhou D, Saint-Mezard P, Wang V, Gao Y, Yin N, et al. Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature. 2005;434:525–529. - PubMed
-
- Kinjo Y, Tupin E, Wu D, Fujio M, Garcia-Navarro R, Benhnia MR, Zajonc DM, Ben-Menachem G, Ainge GD, Painter GF, et al. Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat. Immunol. 2006;7:978–986. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
