

SAP102 mediates synaptic clearance of NMDA receptors
- PMID: 23103165
- PMCID: PMC3513525
- DOI: 10.1016/j.celrep.2012.09.024
SAP102 mediates synaptic clearance of NMDA receptors
Abstract
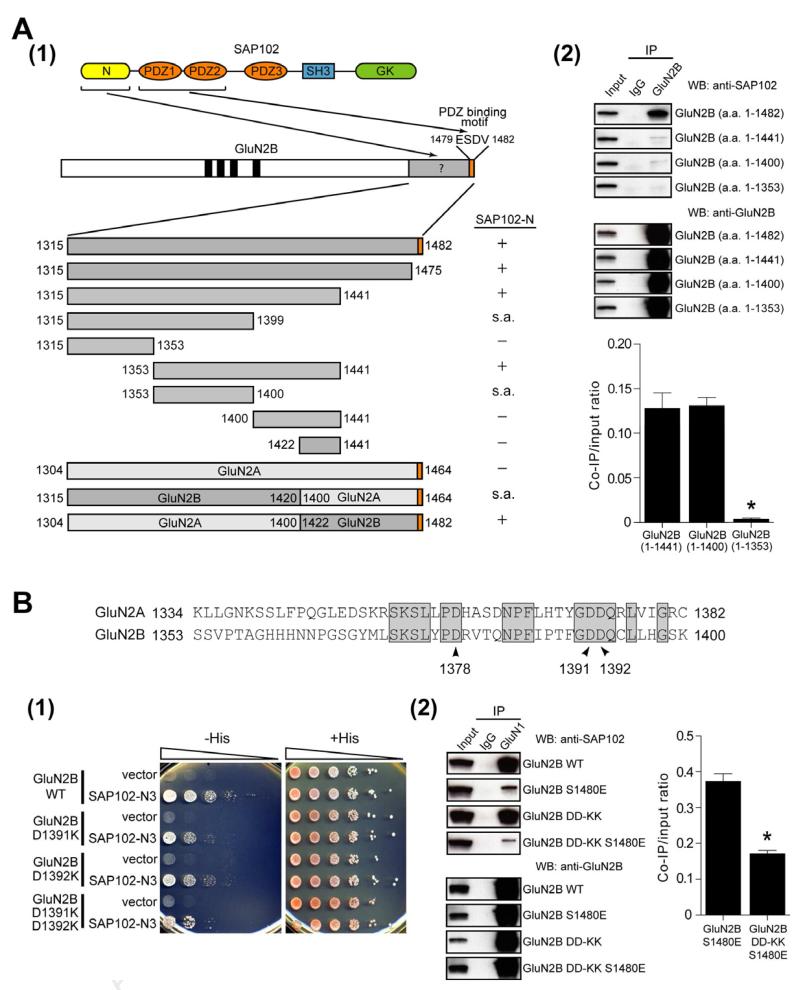
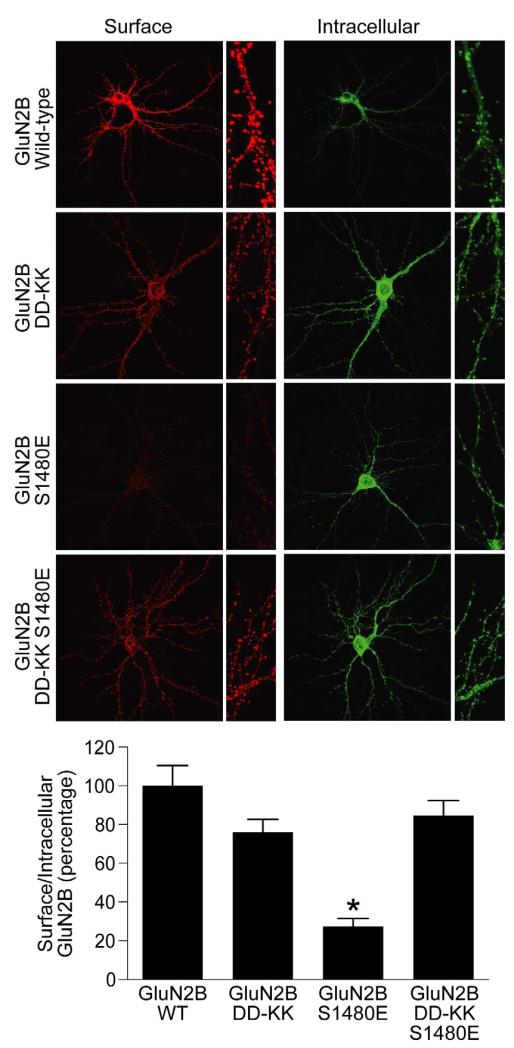
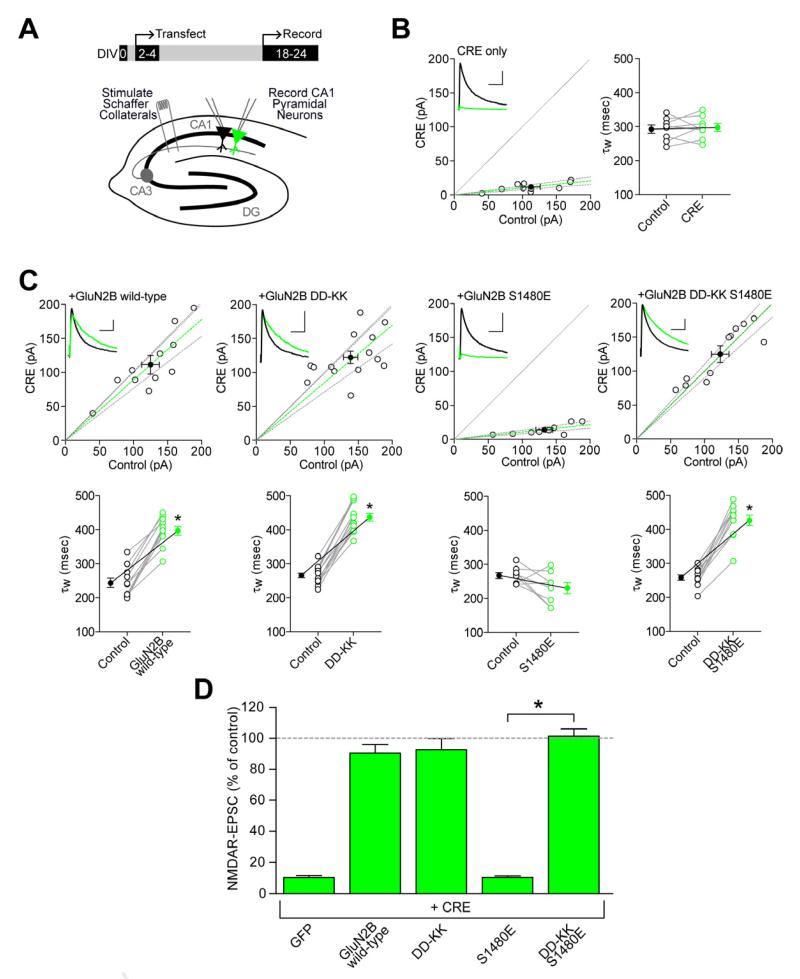
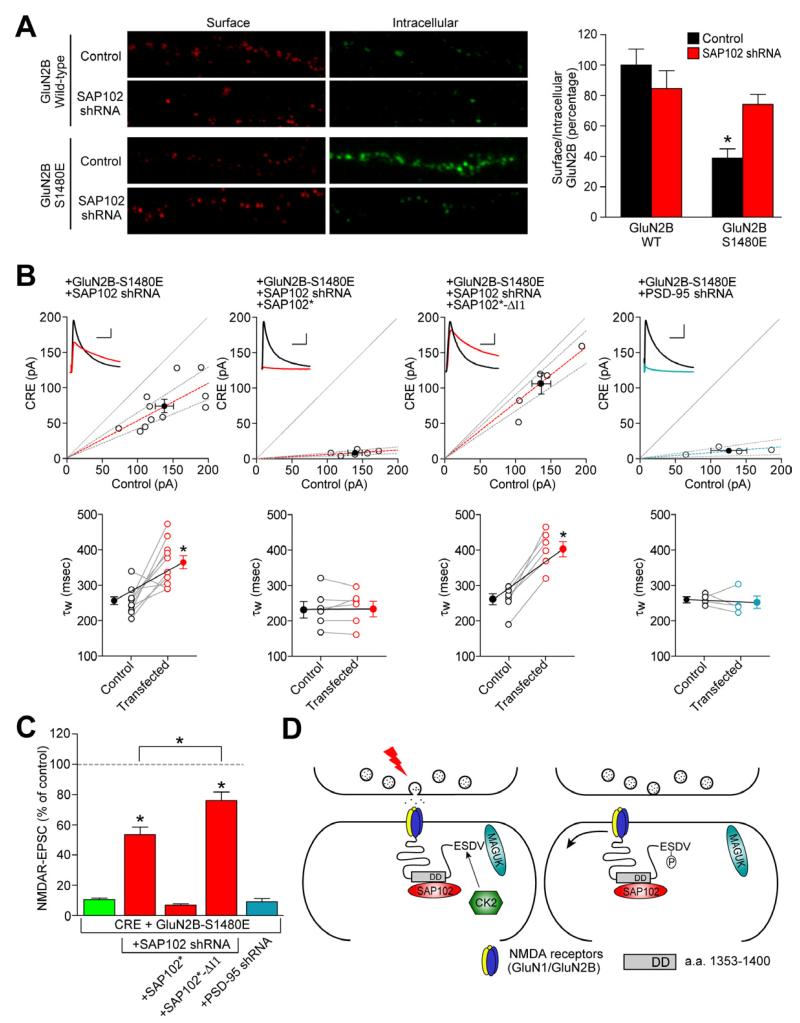
Membrane-associated guanylate kinases (MAGUKs) are the major family of scaffolding proteins at the postsynaptic density. The PSD-MAGUK subfamily, which includes PSD-95, PSD-93, SAP97, and SAP102, is well accepted to be primarily involved in the synaptic anchoring of numerous proteins, including N-methyl-D-aspartate receptors (NMDARs). Notably, the synaptic targeting of NMDARs depends on the binding of the PDZ ligand on the GluN2B subunit to MAGUK PDZ domains, as disruption of this interaction dramatically decreases NMDAR surface and synaptic expression. We recently reported a secondary interaction between SAP102 and GluN2B, in addition to the PDZ interaction. Here, we identify two critical residues on GluN2B responsible for the non-PDZ binding to SAP102. Strikingly, either mutation of these critical residues or knockdown of endogenous SAP102 can rescue the defective surface expression and synaptic localization of PDZ binding-deficient GluN2B. These data reveal an unexpected, nonscaffolding role for SAP102 in the synaptic clearance of GluN2B-containing NMDARs.
Copyright © 2012 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
