

Review
doi: 10.1016/j.heares.2012.09.009.
Epub 2012 Oct 24.
EYA1-SIX1 complex in neurosensory cell fate induction in the mammalian inner ear
Affiliations
- PMID: 23104013
- PMCID: PMC6592049
- DOI: 10.1016/j.heares.2012.09.009
Item in Clipboard
Review
EYA1-SIX1 complex in neurosensory cell fate induction in the mammalian inner ear
Hear Res.
2013 Mar.
Abstract
The phosphatase-transactivator EYA1 interacts with the homeodomain protein SIX1 to form transcriptional activation complexes, which play essential roles in regulating cell proliferation, survival and induction of sensory and neuronal differentiation programs during inner ear development. Mutations of the Eya1 and Six1 genes cause profound developmental auditory defects in mice and humans. The molecular mechanisms and developmental processes controlled by the EYA1 and SIX1 complex in inner ear development and neurosensory fate induction are the focus of this review.
Published by Elsevier B.V.
Figures
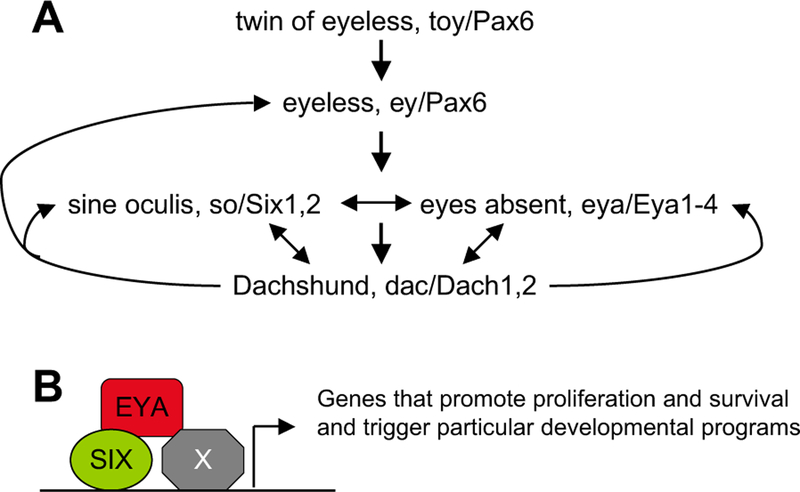
The EYA-SO/SIX regulatory network. (A) The transcriptional hierarchy among the key regulators in the fly eyes. Feedback loops and protein–protein interactions (double-headed arrows) are indicated. (B) EYA and SIX form transcriptional activation complexes to control cell proliferation and survival as well as to trigger particular differentiation programs. EYA–SIX also interact with other proteins (×), which may vary depending on distinct developmental and cellular contexts to modify the specificity of EYA–SIX function.
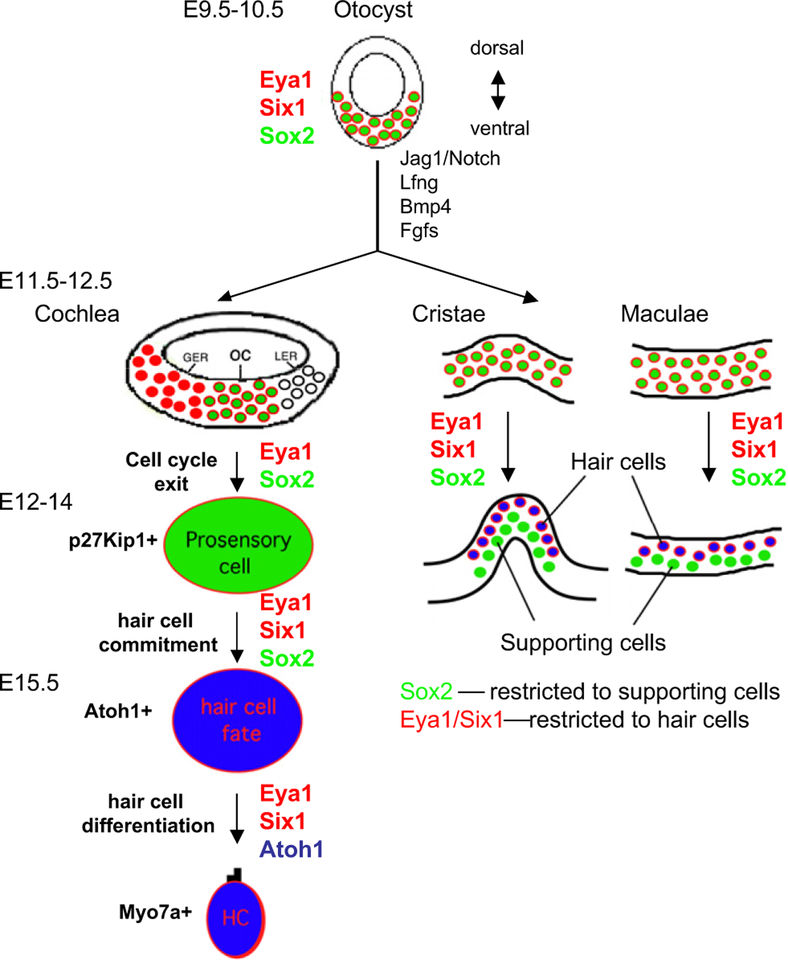
Schematic drawing of Eya1 and Six1 expression domains in relation to Sox2 expression in the otocyst and developing cochlea, cristae and maculae and their possible roles in sensory cell development. In the otocyst, Eya1 (red), Six1 (red) and Sox2 (green) are co-expressed in the ventral otocyst where the prosensory epithelia are formed. These genes may act together to specify a common prosensory area in the otocyst at this early stage by turning on other sensory genes including Jag1, Lfng, Bmp4 and Fgfs, which are not expressed in Eya1- or Six1-null embryos. In the developing cochlea, at E12.5, the floor of cochlear duct will give rise to the organ of Corti (oc), the greater epithelial ridge (GER) and lesser epithelial ridge (LER). Eya1 and Sox2 are coexpressed in the prosenory progenitors of the OC, and this colocalization suggests that both genes may act together to regulate the cell cycle of the prosensory progenitors. Six1 expression disappears in the postmitotic progenitors but reappears in differentiating hair cells in a pattern similar to that of Atoh1. Once the sensory progenitor cells become postmitotic, they express p27Kip1, and at around E13.5–14.5, Atoh1 is activated in the postmitotic progenitors that become committed to hair cell fate and is coexpressed with Eya1 and Six1 in the differentiating hair cells. However, Sox2 expression becomes restricted to supporting cells and border cells in the GER. Based on published data, Eya1, Six1 and Sox2 act together to regulate hair cell commitment by activating Atoh1; however, Eya1 and Six1 but not Sox2 act to maintain or upregulate Atoh1 expression during hair cell differentiation. Similar expression pattern of these genes is found in the sensory cell development of the cristae and maculae.
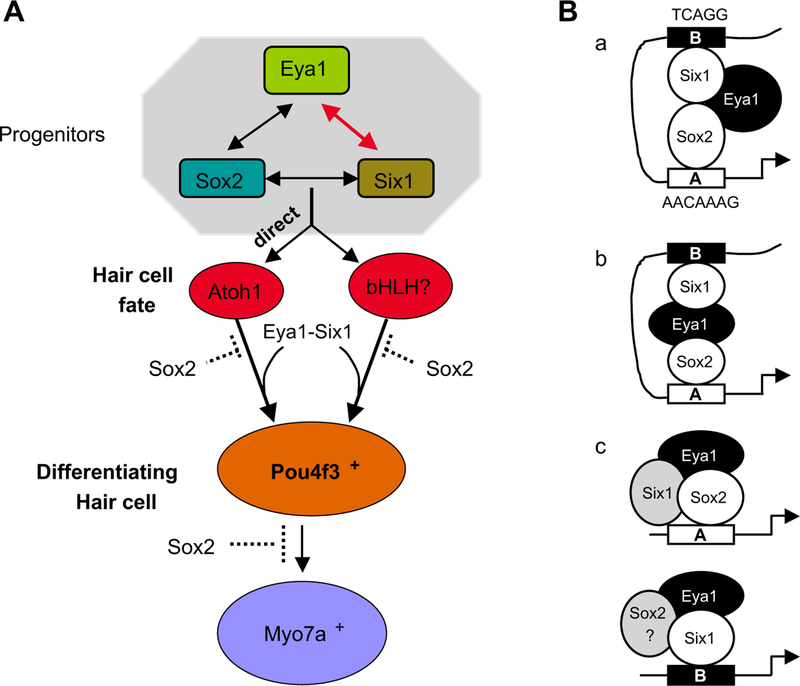
EYA1–SIX1 complex in inducing hair cell fate. (A) Molecular relationships among the key transcription factors for hair cell differentiation. A direct interaction between EYA1/SIX1/SOX2 proteins coordinately regulates Atoh1 expression, and that POU4F3 is a common downstream factor of the Atoh1-dependent and -independent pathways. Dashed lines indicate that SOX2 may repress Pou4f3 or downstream factors of Pou4f3 to inhibit hair cell differentiation. (B) Possible mechanisms for Atoh1 activation by Sox2/Eya1/Six1. EYA1/SIX1 in collaboration with SOX2 activity in prosensory progenitors can induce Atoh1 activation via direct binding to the Sox and Six binding sites within enhancer A and B, respectively. These three factors may directly interact (model a), or EYA1 may bridge SIX1 and SOX2 (model b). These three factors may also form an active complex to regulate Atoh1 activation via enhancer A (model c), whereas EYA1/SIX1 efficiently upregulate Atoh1 via enhancer B. Question mark indicates that the involvement of the factor is unclear.
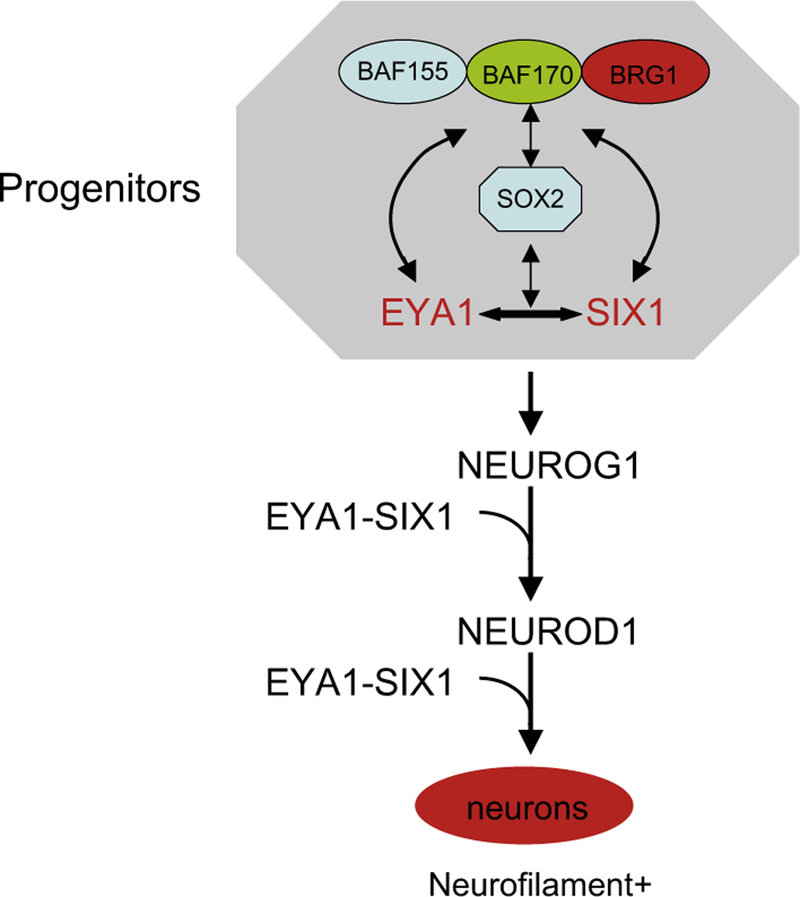
Possible mechanism for Neurog1 and Neurod1 activation by Eya1/Six1. EYA1/SIX1 in collaboration with the SWI/SNF complex in progenitor cells induce Neurog1—Neurod1 activation. Eya1/Six1 also interact with Neurog1 to regulate Neurod1 activation. In addition, Eya1/Six1 interact with Neurod1 to regulate neuronal differentiation. Sox2 works cooperatively with Eya1/Six1 or the SWI/SNF complex to promote neuronal differentiation. These factors are physically associated.
Similar articles
-
Eya1 is required for lineage-specific differentiation, but not for cell survival in the zebrafish adenohypophysis.Dev Biol. 2006 Apr 1;292(1):189-204. doi: 10.1016/j.ydbio.2005.12.036. Epub 2006 Feb 3. Dev Biol. 2006. PMID: 16458879
-
Eya1 regulates the growth of otic epithelium and interacts with Pax2 during the development of all sensory areas in the inner ear.Dev Biol. 2006 Oct 15;298(2):430-41. doi: 10.1016/j.ydbio.2006.06.049. Epub 2006 Jul 7. Dev Biol. 2006. PMID: 16916509 Free PMC article.
-
Eya1-Six1 interaction is sufficient to induce hair cell fate in the cochlea by activating Atoh1 expression in cooperation with Sox2.Dev Cell. 2012 Feb 14;22(2):377-90. doi: 10.1016/j.devcel.2011.12.006. Dev Cell. 2012. PMID: 22340499 Free PMC article.
-
The EYA-SO/SIX complex in development and disease.Pediatr Nephrol. 2013 Jun;28(6):843-54. doi: 10.1007/s00467-012-2246-1. Epub 2012 Jul 19. Pediatr Nephrol. 2013. PMID: 22806561 Free PMC article. Review.
-
Transcription factors that control inner ear development and their potential for transdifferentiation and reprogramming.Hear Res. 2013 Mar;297:84-90. doi: 10.1016/j.heares.2012.11.001. Epub 2012 Nov 14. Hear Res. 2013. PMID: 23159917 Review.
Cited by
-
Bioelectric signalling via potassium channels: a mechanism for craniofacial dysmorphogenesis in KCNJ2-associated Andersen-Tawil Syndrome.J Physiol. 2016 Jun 15;594(12):3245-70. doi: 10.1113/JP271930. Epub 2016 Apr 13. J Physiol. 2016. PMID: 26864374 Free PMC article.
-
Open chromatin dynamics in prosensory cells of the embryonic mouse cochlea.Sci Rep. 2019 Jun 21;9(1):9060. doi: 10.1038/s41598-019-45515-2. Sci Rep. 2019. PMID: 31227770 Free PMC article.
-
The transcriptional coactivator Eya1 exerts transcriptional repressive activity by interacting with REST corepressors and REST-binding sequences to maintain nephron progenitor identity.Nucleic Acids Res. 2022 Oct 14;50(18):10343-10359. doi: 10.1093/nar/gkac760. Nucleic Acids Res. 2022. PMID: 36130284 Free PMC article.
-
EYA1's Conformation Specificity in Dephosphorylating Phosphothreonine in Myc and Its Activity on Myc Stabilization in Breast Cancer.Mol Cell Biol. 2016 Dec 19;37(1):e00499-16. doi: 10.1128/MCB.00499-16. Print 2017 Jan 1. Mol Cell Biol. 2016. PMID: 27795300 Free PMC article.
-
Multiple Functions of the Eya Phosphotyrosine Phosphatase.Mol Cell Biol. 2015 Dec 14;36(5):668-77. doi: 10.1128/MCB.00976-15. Mol Cell Biol. 2015. PMID: 26667035 Free PMC article. Review.
References
-
- Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, Levi-Acobas F, Cruaud C, Le Merrer M, Mathieu M, Konig R, Vigneron J, Weissenbach J, Petit C, Weil D, 1997a. Clustering of mutations responsible for branchio-oto-renal (BOR) syndrome in the eyes absent homologous region (eyaHR) of EYA1. Hum. Mol. Genet 6, 2247–2255. - PubMed
-
- Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, Weil D, Cruaud C, Sahly I, Leibovici M, Bitner-Glindzicz M, Francis M, Lacombe D, Vigneron J, Charachon R, Boven K, Bedbeder P, Van Regemorter N, Weissenbach J, Petit C, 1997b. A human homologue of the Drosophila eyes absent gene underlies branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nat. Genet 15, 157–164. - PubMed
-
- Barald KF, Kelley MW, 2004. From placode to polarization: new tunes in inner ear development. Development 131, 4119–4130. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
