

Mechanism of repair of 5'-topoisomerase II-DNA adducts by mammalian tyrosyl-DNA phosphodiesterase 2
- PMID: 23104055
- PMCID: PMC3529160
- DOI: 10.1038/nsmb.2418
Mechanism of repair of 5'-topoisomerase II-DNA adducts by mammalian tyrosyl-DNA phosphodiesterase 2
Abstract
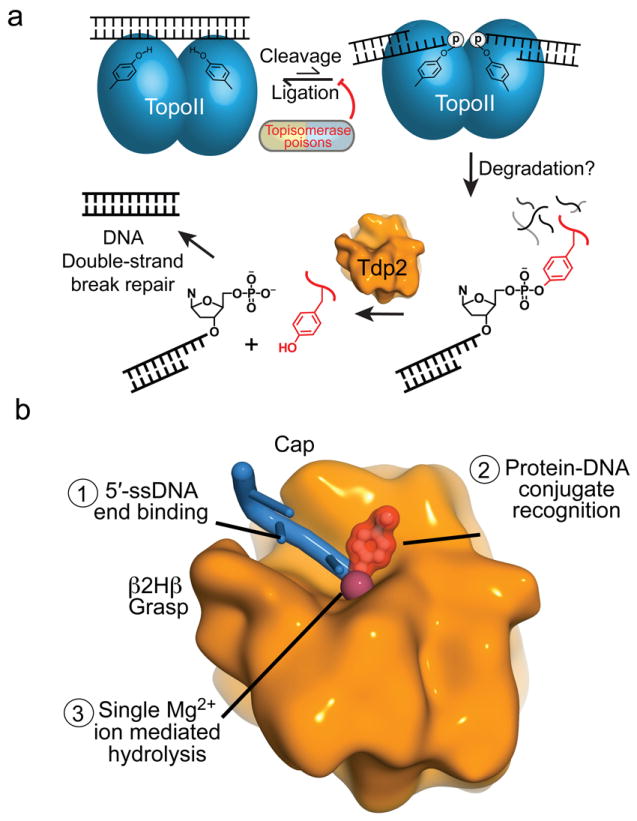
The topoisomerase II (topo II) DNA incision-and-ligation cycle can be poisoned (for example following treatment with cancer chemotherapeutics) to generate cytotoxic DNA double-strand breaks (DSBs) with topo II covalently conjugated to DNA. Tyrosyl-DNA phosphodiesterase 2 (Tdp2) protects genomic integrity by reversing 5'-phosphotyrosyl-linked topo II-DNA adducts. Here, X-ray structures of mouse Tdp2-DNA complexes reveal that Tdp2 β-2-helix-β DNA damage-binding 'grasp', helical 'cap' and DNA lesion-binding elements fuse to form an elongated protein-DNA conjugate substrate-interaction groove. The Tdp2 DNA-binding surface is highly tailored for engagement of 5'-adducted single-stranded DNA ends and restricts nonspecific endonucleolytic or exonucleolytic processing. Structural, mutational and functional analyses support a single-metal ion catalytic mechanism for the exonuclease-endonuclease-phosphatase (EEP) nuclease superfamily and establish a molecular framework for targeted small-molecule blockade of Tdp2-mediated resistance to anticancer topoisomerase drugs.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
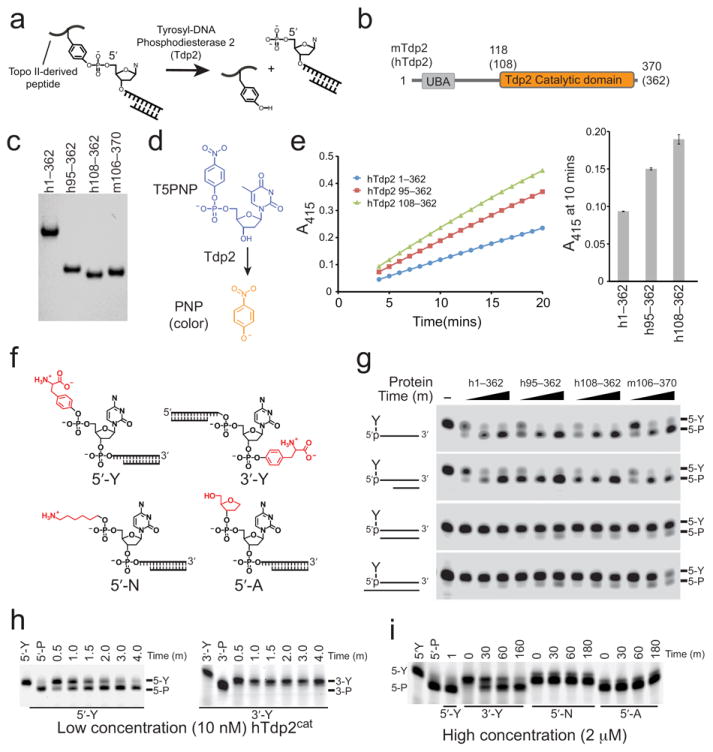
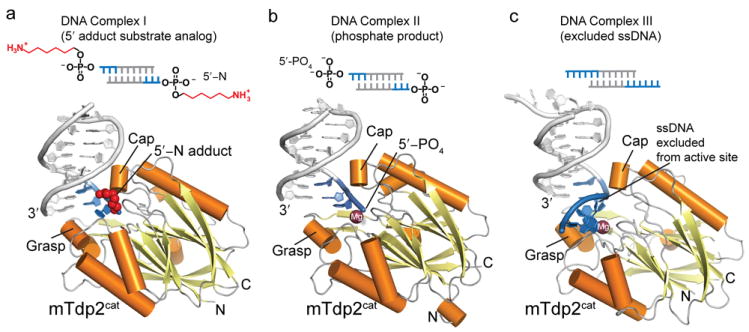
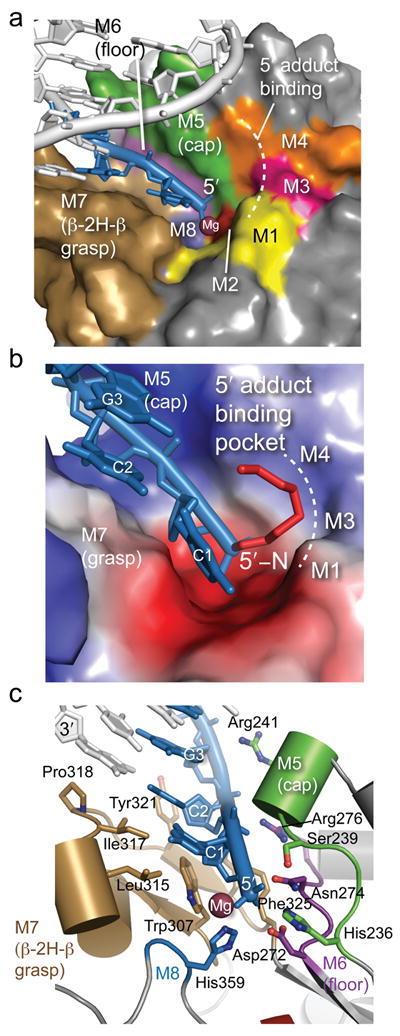
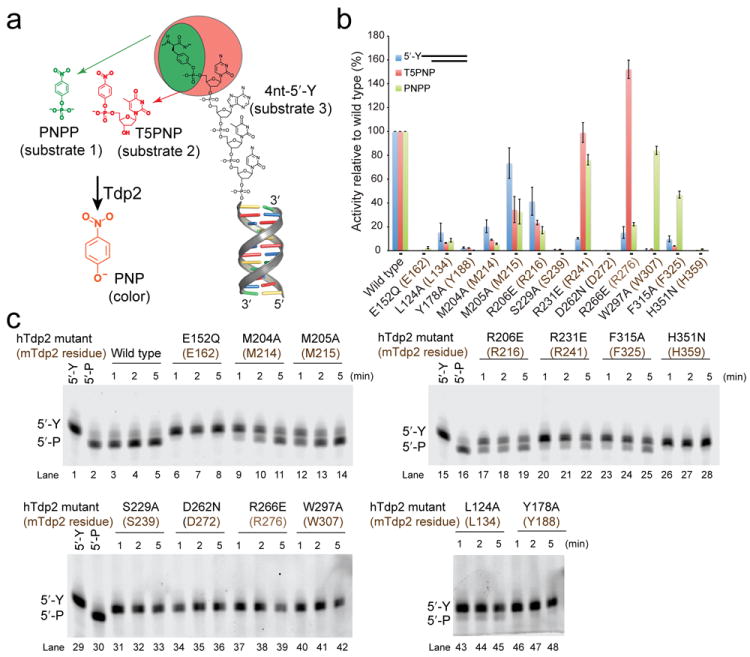
Comment in
-
Tyrosyl DNA phosphodiesterase 2, an enzyme fit for purpose.Nat Struct Mol Biol. 2012 Dec;19(12):1212-3. doi: 10.1038/nsmb.2455. Nat Struct Mol Biol. 2012. PMID: 23211766 No abstract available.
References
-
- Wilstermann AM, Osheroff N. Base excision repair intermediates as topoisomerase II poisons. J Biol Chem. 2001;276:46290–6. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
