

Structural basis for recognition of 5'-phosphotyrosine adducts by Tdp2
- PMID: 23104058
- PMCID: PMC3515695
- DOI: 10.1038/nsmb.2423
Structural basis for recognition of 5'-phosphotyrosine adducts by Tdp2
Abstract
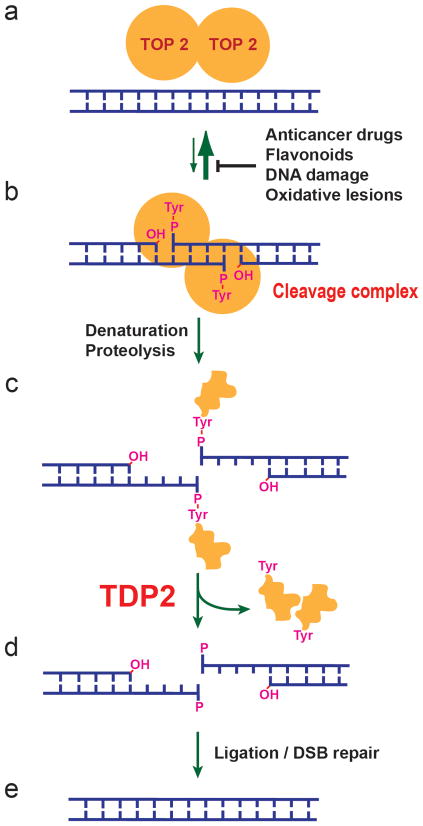
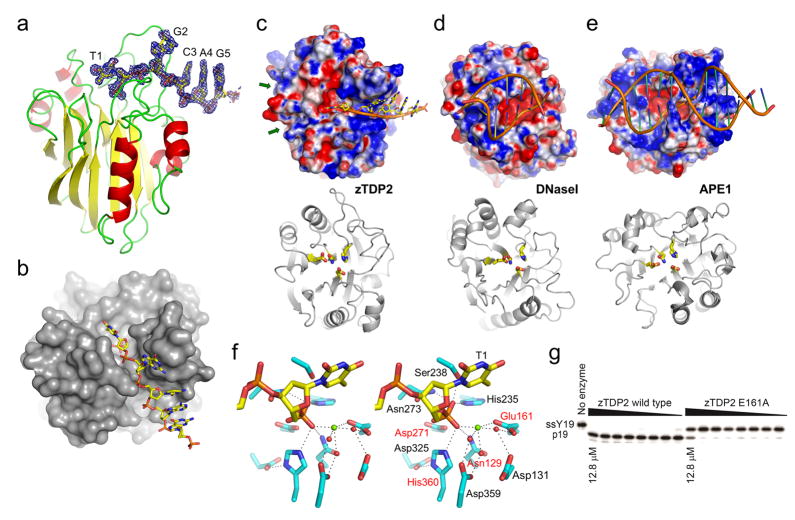
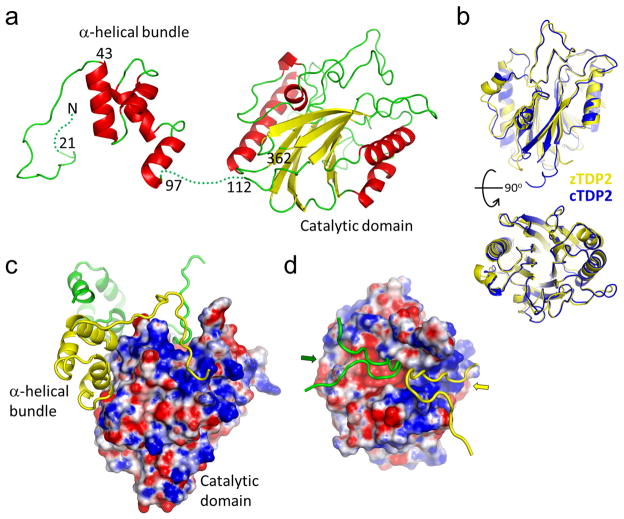
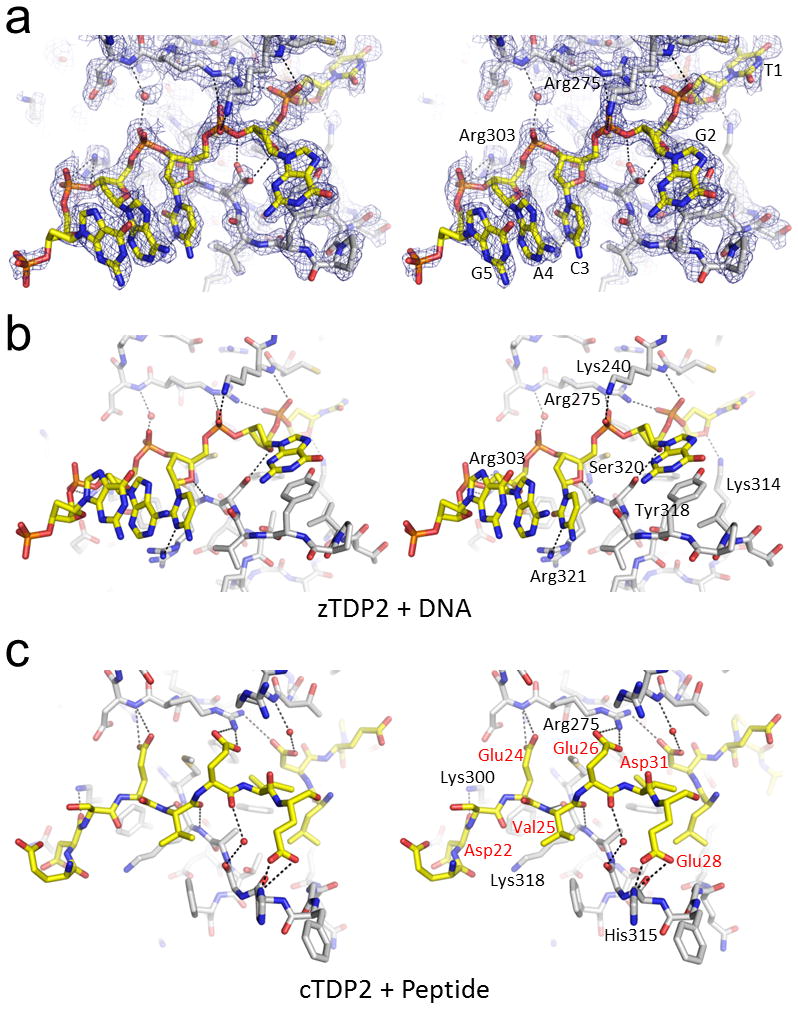
The DNA-repair enzyme Tdp2 resolves 5'-phosphotyrosyl DNA adducts and mediates resistance to anticancer drugs that target covalent topoisomerase-DNA complexes. Tdp2 also participates in key signaling pathways during development and tumorigenesis and cleaves a protein-RNA linkage during picornavirus replication. The crystal structure of zebrafish Tdp2 bound to DNA reveals a deep, narrow basic groove that selectively accommodates the 5' end of single-stranded DNA in a stretched conformation. The crystal structure of the full-length Caenorhabditis elegans Tdp2 shows that this groove can also accommodate an acidic peptide stretch in vitro, with glutamate and aspartate side chains occupying the DNA backbone phosphate-binding sites. This extensive molecular mimicry suggests a potential mechanism for autoregulation and interaction of Tdp2 with phosphorylated proteins in signaling. Our study provides a framework to interrogate functions of Tdp2 and develop inhibitors for chemotherapeutic and antiviral applications.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Comment in
-
Tyrosyl DNA phosphodiesterase 2, an enzyme fit for purpose.Nat Struct Mol Biol. 2012 Dec;19(12):1212-3. doi: 10.1038/nsmb.2455. Nat Struct Mol Biol. 2012. PMID: 23211766 No abstract available.
References
-
- Wang JC. Cellular roles of DNA topoisomerases: a molecular perspective. Nat Rev Mol Cell Biol. 2002;3:430–440. - PubMed
-
- Champoux JJ. DNA topoisomerases: structure, function, and mechanism. Annu Rev Biochem. 2001;70:369–413. - PubMed
-
- Corbett KD, Berger JM. Structure, molecular mechanisms, and evolutionary relationships in DNA topoisomerases. Annu Rev Biophys Biomol Struct. 2004;33:95–118. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- Z01 BC006150/ImNIH/Intramural NIH HHS/United States
- R21 AI087098/AI/NIAID NIH HHS/United States
- P41 RR015301/RR/NCRR NIH HHS/United States
- P01 CA092584/CA/NCI NIH HHS/United States
- R01 GM046312/GM/NIGMS NIH HHS/United States
- AI087098/AI/NIAID NIH HHS/United States
- R33 AI087098/AI/NIAID NIH HHS/United States
- Z01 BC 006150-19/BC/NCI NIH HHS/United States
- R01GM105404/GM/NIGMS NIH HHS/United States
- GM095558/GM/NIGMS NIH HHS/United States
- GM046312/GM/NIGMS NIH HHS/United States
- R01 GM105404/GM/NIGMS NIH HHS/United States
- RR15301/RR/NCRR NIH HHS/United States
- R01 GM095558/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
