

Liver retinol transporter and receptor for serum retinol-binding protein (RBP4)
- PMID: 23105095
- PMCID: PMC3543007
- DOI: 10.1074/jbc.M112.369132
Liver retinol transporter and receptor for serum retinol-binding protein (RBP4)
Abstract
Vitamin A (retinol) is absorbed in the small intestine, stored in liver, and secreted into circulation bound to serum retinol-binding protein (RBP4). Circulating retinol may be taken up by extrahepatic tissues or recycled back to liver multiple times before it is finally metabolized or degraded. Liver exhibits high affinity binding sites for RBP4, but specific receptors have not been identified. The only known high affinity receptor for RBP4, Stra6, is not expressed in the liver. Here we report discovery of RBP4 receptor-2 (RBPR2), a novel retinol transporter expressed primarily in liver and intestine and induced in adipose tissue of obese mice. RBPR2 is structurally related to Stra6 and highly conserved in vertebrates, including humans. Expression of RBPR2 in cultured cells confers high affinity RBP4 binding and retinol transport, and RBPR2 knockdown reduces RBP4 binding/retinol transport. RBPR2 expression is suppressed by retinol and retinoic acid and correlates inversely with liver retinol stores in vivo. We conclude that RBPR2 is a novel retinol transporter that potentially regulates retinol homeostasis in liver and other tissues. In addition, expression of RBPR2 in liver and fat suggests a possible role in mediating established metabolic actions of RBP4 in those tissues.
Figures



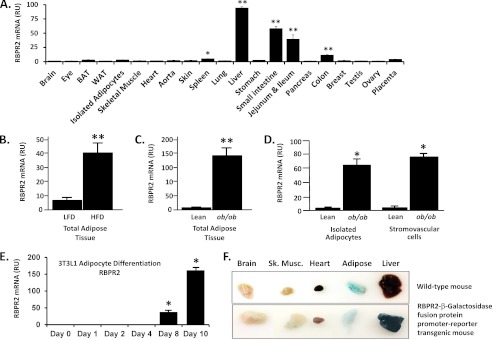
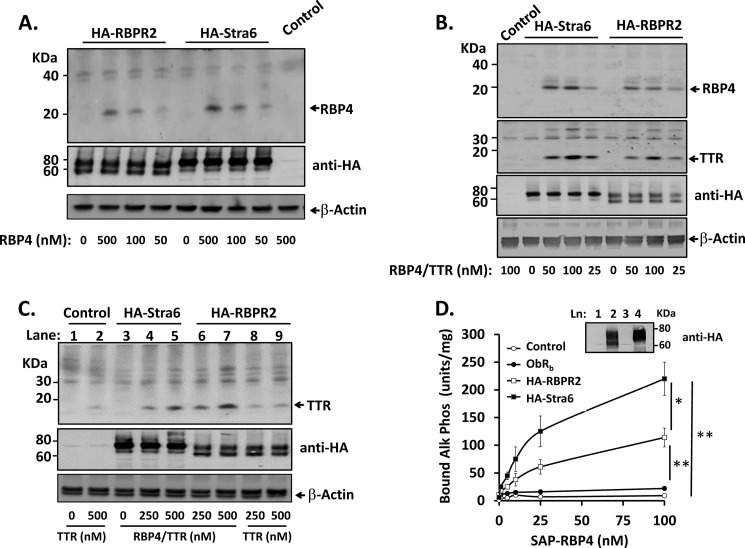




References
-
- Norum K. R., Blomhoff R. (1992) McCollum Award Lecture, 1992. Vitamin A absorption, transport, cellular uptake, and storage. Am. J. Clin. Nutr. 56, 735–744 - PubMed
-
- Chambon P. (1996) A decade of molecular biology of retinoic acid receptors. FASEB J. 10, 940–954 - PubMed
-
- McGrane M. M. (2007) Vitamin A regulation of gene expression. Molecular mechanism of a prototype gene. J. Nutr. Biochem. 18, 497–508 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
